- Trang chủ
- Sách y học
- Bài giảng miễn dịch
- Cấu trúc phân tử của kháng thể
Cấu trúc phân tử của kháng thể
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Các nghiên cứu trước đây về kháng thể chủ yếu là lấy kháng thể từ máu các cá thể được gây miễn dịch rồi sau đó tinh chế để khảo sát. Nhưng cách này thì không thể dùng để xác định cấu trúc của kháng thể một cách chính xác, bởi vì huyết thanh chứa quá nhiều loại kháng thể được sản xuất bởi nhiều clôn tế bào B đáp ứng với nhiều quyết định kháng nguyên (epitốp) khác nhau (và được gọi là kháng thể đa clôn). Có hai phát hiện đột phá đã tạo điều kiện để có thể kháo sát cấu trúc kháng thể một cách chính xác. Phát hiện thứ nhất là bệnh nhân đa u tủy có một khối u tương bào tiết ra kháng thể đơn clôn với lượng đủ lớn trong máu và nước tiểu để có thể xác định tính chất hóa sinh của chúng. Phát hiện thứ hai, quan trọng hơn, là kỹ thuật tạo ra kháng thể đơn clôn được Georges Kohler và Cesar Milstein mô tả năm 1975. Họ đã tìm được cách làm cho tế bào sản xuất kháng thể bất tử bằng kỹ thuật tạo “u lai” (hybridoma); những u lai này cho phép sản xuất kháng thể chống lại chỉ một tính đặc hiệu kháng nguyên đã định trước. Việc tạo ra được quần thể tế bào sản xuất kháng thể thuần nhất này đã giúp cho các nhà nghiên cứu xác định được trình tự acid amin của từng phân tử kháng thể.
Đặc điểm chung của cấu trúc kháng thể
Các glycoprotein huyết tương hoặc huyết thanh trước đây thường được phân chia theo tính chất hòa tan của chúng thành albumin và globulin và phân chia sâu hơn dựa theo tính điện di của chúng. Hầu hết kháng thể được tìm thấy trong thành phần điện di nhanh thứ ba của globulin và được gọi là gamma globulin (gamma là chữ cái thứ ba trong mẫu tự Hy Lạp). Một tên thường gọi nữa của kháng thể là immunoglobulin (globulin miễn dịch), viết tắt là Ig. Trong quyển sách này hai thuật ngữ immunoglobulin và kháng thể thường được dùng với cùng một ý nghĩa.
Tất cả mọi phân tử kháng thể đều có chung một cấu trúc cơ bản giống nhau, và sự khác biệt chỉ thấy ở vị trí kết hợp kháng nguyên. Sự khác biệt của vùng kết hợp kháng nguyên giúp cho kháng thể liên kết dễ dàng với kháng nguyên về mặt cấu trúc. Trong mỗi cá thể có khoảng từ 107 đến 109 phân tử kháng thể khác nhau mang những vùng kết hợp kháng nguyên khác nhau. Các chức năng hiệu quả và tính chất hóa lý của kháng thể thường do phần không liên quan đến kháng nguyên đảm trách và những phần này thường không khác nhau mấy giữa các kháng thể.
Mỗi phân tử kháng thể có một cấu trúc lõi đối xứng được cấu tạo bởi hai chuỗi nặng và hai chuỗi nhẹ giống hệt nhau (Hình 3.1). Mỗi chuỗi nhẹ có trọng lượng phân tử khoảng 24 kD, và mỗi chuỗi nặng khoảng 55-70 kD. Một chuỗi nhẹ âæåüc liên kết đồng hóa trị với một chuỗi nặng bằng một cầu disulphua và các chuỗi nặng cũng nối với nhau bằng những cầu disulphua. Cả chuỗi nhẹ và nặng đều chứa những chuỗi đơn vị đồng nhất và lặp đi lặp lại. Mỗi đơn vị có độ dài 110 acid amin cuộn lại một cách độc lập thành những búi gọi là lĩnh vực Ig (Ig domain). Một lĩnh vực Ig chứa hai tấm gấp β, mỗi lớp có 3-5 chuỗi polypeptid. Có nhiều protein quan trọng khác trong hệ thống miễn dịch cũng chứa những lĩnh vực có cấu tạo mô-típ tấm gấp tương tự như vậy và có các chuỗi acid amin giống như Ig. Tất cả những phân tử chứa cấu tạo theo mô-típ này được đặt chung một tên là siêu họ Ig (Ig superfamily), và tất cả những gen mã hóa cho các lĩnh vực Ig của những phân tử này được cho là đều tiến hóa từ một gen tổ tiên.
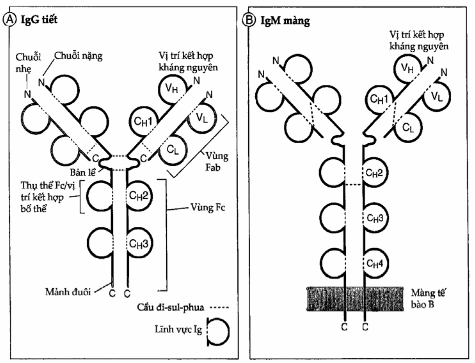
Hình. Cấu trúc của một phân tử kháng thể.
A. Sơ đồ cấu trúc của một phân tử IgG. Diện kết hợp kháng nguyên được tạo bởi lĩnh vực thay đổi của chuỗi nhẹ (VL) và chuỗi nặng (VH) nằm cạnh nhau. Các vùng C của chuỗi nặng tận cùng tại phần đuôi.
B. Sơ đồ cấu trúc của phân tử IgM gắn trên màng tế bào lymphô B. IgM có nhiều hơn IgG một lĩnh vực vùng C chuỗi nặng (CH) và tận cùng bằng một đoạn C cắm xuyên qua màng tế bào.
C. Cấu trúc của IgG người được vẽ nhờ phép chụp tinh thể bằng tia X. Trong sơ đồ này chuỗi nặng được vẽ bằng màu xanh nước biển và đỏ, còn chuỗi nhẹ có màu lục.
Cả chuỗi nặng và nhẹ đều mang một đầu tận cùng amin luôn thay đổi (gọi là vùng biến đổi hay vùng V) tham gia vào việc nhận diện kháng nguyên và một đầu tận cùng cacboxyl hằng định (gọi là vùng hằng định hay vùng C); vùng hằng định của chuỗi nặng làm trung gian cho các chức năng hiệu quả. Trên chuỗi nặng, vùng V chỉ có 1 lĩnh vực Ig, nhưng vùng C thì có 3-4 lĩnh vực Ig. Mỗi chuỗi nhẹ chỉ cấu tạo bởi 1 lĩnh vực Ig vùng V và 1 lĩnh vực Ig vùng C. Vùng biến đổi của mỗi phân tử kháng thể giúp phân biệt kháng thể được sản xuất bởi một clôn tế bào B này với kháng thể được sản xuất bởi một clôn tế bào B khác. Vùng biến đổi của chuỗi nặng (được gọi là VH) nằm sát bên cạnh vùng V của chuỗi nhẹ (được gọi là VL) để tạo nên diện kết hợp kháng nguyên (Hình 3.1). Bởi vì đơn vị cấu tạo cơ bản của mỗi phân tử kháng thể chứa 2 chuỗi nặng và 2 chuỗi nhẹ nên nó có 2 diện kết hợp kháng nguyên. Các lĩnh vực của vùng C tách biệt với diện kết hợp kháng nguyên và không tham gia vào quá trình nhận diện kháng nguyên. Các vùng C của chuỗi nặng có khả năng tương tác với các phân tử và tế bào hiệu quả khác của hệ miễn dịch nên do đó đảm trách hầu hết các chức năng sinh học của kháng thể. Ngoaìi ra, đầu tận cùng cacboxyl của các chuỗi nặng có tác dụng gắn các phân tử kháng thể màng vào màng tế bào B. Vùng C của chuỗi nhẹ không tham gia vào các chức năng hiệu quả và cũng không dính vào màng tế bào.
Đặc điểm cấu trúc của vùng biến đổi và mối liên quan với sự liên kết kháng nguyên
Mỗi một kháng thể thường mang 3 đoạn ngắn acid amin ở vùng V (của cả chuỗi nặng và chuỗi nhẹ) rất riêng cho kháng thể đó và được gọi là vùng siêu biến. Những vùng siêu biến này có độ dài khoảng 10 acid amin và được giữ yên tại vị trí của nó chuỗi peptid tạo nên lĩnh vực của vùng V. Trong mỗi phân tử kháng thể, 3 vùng siêu biến của vùng VL và 3 vùng siêu biến của vùng VH được kéo lại với nhau để tạo nên một cấu trúc không gian ba chiều, đó là diện kết hợp kháng nguyên. Bởi vì cấu tạo này được tạo ra sao cho khớp với cấu trúc không gian ba chiều của phân tử kháng nguyên tương ứng nên những vùng siêu biến này còn được gọi là vùng xác định tính bổ khuyết (complementarity-determined regions, CDR). Trên mỗi chuỗi acid amin của VL hoặc VH, các vùng này được đặt tên là CDR1, CDR2 và CDR3. Vùng CDR3 của cả VL và HL là vùng có độ biến đổi cao nhất trong số 3 vùng vừa kể. Có những cơ chế di truyền tạo ra tính đa dạng cao hơn ở CDR3 so với CDR1 và CDR2. Nghiên cứu hình ảnh tinh thể của kháng thể cho thấy các CDR tạo ra những cung hóa học đưa ra bề ngoài kháng thể để sẵn sàng tương tác với kháng nguyên. Sự khác biệt về trình tự acid amin trong các CDR của các phân tử kháng thể khác nhau tạo ra cấu trúc hóa học riêng biệt cho chúng và được thể hiện ra bề mặt của kháng thể để tạo nên tính đặc hiệu đối với kháng nguyên.
Đặc điểm cấu trúc của vùng hằng định và mối liên quan với chức năng hiệu quả
Các phân tử kháng thể có thể được chia thành nhiều lớp khác nhau dựa trên sự khác biệt về cấu trúc vùng C của chuỗi nặng. Các lớp kháng thể còn được gọi là isotyp và gồm có 5 lớp được đặt tên là: IgA, IgD, IgE, IgG, và IgM. Ở người, các isotyp IgA và IgG có thể được chia thành các tiểu lớp và đặt tên là: IgA1, IgA2, IgG1, IgG2, IgG3, IgG4. (ở chuột nhắt, loài động vật thường được dùng trong nghiên cứu miễn dịch học, thì lớp IgG được chia thành các tiểu lớp IgG1, IgG2a, IgG2b, và IgG3.) Vùng C của chuỗi nặng của tất cả các phân tử kháng thể trong một lớp hoặc tiểu lớp đều có chung một trình tự acid amin. Trình tự này khác với trình tự của lớp khác hay tiểu lớp khác. Các chuỗi nặng được đặt tên theo chữ cái Hy Lạp tương ứng với isotyp kháng thể: IgA1 chứa chuỗi nặng α1; IgA2 chứa α2; IgD chứa δ; IgE chứa ε; IgG1 chứa γ1; IgG2 chứa γ2; IgG3 chứa γ3; IgG4 chứa γ4 và IgM chứa μ. Trong kháng thể của IgM và IgE người, vùng C chứa 4 lĩnh vực Ig nối tiếp nhau. Vùng C của IgG, IgA, và IgD chỉ chứa 3 lĩnh vực Ig. Những lĩnh vực này được ký hiệu là CH và được đánh số từ đầu tận cùng amin đến đầu tận cùng cacboxyl (ví dụ CH1, CH2, .v.v.). Trong mỗi isotyp, các vùng này được ký hiệu cụ thể hơn (ví dụ Cγ1, Cγ2 trong IgG). Kháng thể có thể có tác dụng như một kháng nguyên khi nó được đưa vào một cơ thể lạ và kích thích cơ thể đó tạo kháng - kháng thể. Bằng cách gây miễn dịch một con vật bằng kháng thể lấy từ một con vật khác loài, chúng ta có thể sản xuất ra các kháng - kháng thể đặc hiệu cho một lớp hoặc tiểu lớp Ig, và kháng thể kiểu này thường được dùng thường xuyên trong các nghiên cứu lâm sàng hoặc thực nghiệm về đáp ứng miễn dịch dịch thể.
Những lớp và tiểu lớp kháng thể khác nhau sẽ tạo nên những chức năng hiệu quả khác nhau. Nguyên nhân là vì hầu hết các chức năng hiệu quả đều qua trung gian của sự kết hợp vùng C chuỗi nặng vào thụ thể của nó lên các loại tế bào như đại thực bào, tế bào NK, tế bào mast, hoặc kết hợp với các loại protein huyết tương khác nhau như protein bổ thể. Các lớp và tiểu lớp kháng thể khác nhau ở vùng C nên cũng khác nhau về chức năng hiệu quả.
Chuỗi nhẹ của kháng thể có hai lớp và được gọi là κ và λ phân biệt nhờ vùng hằng định tận cùng bằng cacboxyl. Một phân tử kháng thể có thể có hoặc là 2 chuỗi nhẹ κ hoặc là 2 chuỗi nhẹ λ, và không bao giờ có mỗi loại một chuỗi. Trình tự acid amin của vùng C chuỗi nhẹ κ (Cκ) khác với trình tự đó của vùng C chuỗi nhẹ λ (Cλ), nhưng trình tự Cκ của tất cả các phân tử kháng thể đều giống hệt nhau, và trình tự Cλ cũng vậy. Mặc dù có khác nhau về trình tự acid amin nhưng Cκ và Cλ hoàn toàn đồng nhất về mặt cấu trúc, và đều gập vào trong cùng một lĩnh vực Ig. Ở người, khoảng 60% phân tử kháng thể có chuỗi nhẹ κ và 40% có chuỗi λ. Người ta tìm thấy tỉ lệ này thay đổi rất rõ trên bệnh nhân u tế bào B đơn clôn bởi vì các clôn tế bào ung thư đã sản xuất những phân tử kháng thể với cùng một lớp chuỗi nhẹ mà thôi. Và người ta đã dùng tỉ lệ tế bào mang κ và tế bào mang λ để chẩn đoán u lymphô tế bào B. Ở chuột, lượng kháng thể chứa κ nhiều gấp 10 lần lượng kháng thể chứa λ. Khác với lớp chuỗi nặng, các lớp chuỗi nhẹ không tạo ra điều gì khác nhau về chức năng của kháng thể.
Kháng thể có thể xuất hiện dưới dạng chất tiết (dạng tiết) hoặc nằm trên màng tế bào (dạng màng). Dạng kháng thể này khác với kháng thể huyết thanh về trình tự acid amin ở đầu tận cùng cacboxyl của vùng C chuỗi nặng. Dạng tiết gặp ở trong máu và các dịch ngoại bào. Đối với dạng này, trình tự acid amin của vùng CH được tận cùng bằng các acid amin ưa nước và tích điện. Đây là cấu trúc đặc trưng cho các protein gắn trên màng tế bào.
Các kháng thể IgG và IgE được tiết ra dưới dạng đơn phân tử tức ở dạng đơn vị kháng thể cơ bản (bao gồm 2 chuỗi nặng và 2 chuỗi nhẹ). Ngược lại, các kháng thể IgM và IgA thì thường tạo nên những phức hợp đa phân tử tức bao gồm từ 4 chuỗi nặng và 4 chuỗi nhẹ trở lên. Các kháng thể đa phân tử IgM và IgA còn chứa một polypeptid phụ với trọng lượng phân tử 15 kD gọi là chuỗi J (liên kết), gắn với các phân tử kháng thể bởi các cầu nối disulphua và có chức năng làm bền vững các phức hợp đa phân tử này.
Sự tổng hợp và biểu hiện của phân tử Ig
Các chuỗi nặng và nhẹ của Ig được tổng hợp trong các ribosom nằm trên bề mặt của hệ lưới nội bào. Sự kết hợp đồng hóa trị của các chuỗi nặng và các chuỗi nhẹ được thực hiện nhờ các cầu nối disulphua và cũng xảy ra trong hệ lưới nội bào. Sau khi được hình thành phân tử Ig được chuyển đến thể Golgi từ đó chuyển ra màng tế bào trong các nang nhỏ và sau đó được tiết ra khỏi tế bào qua một quá trình gọi là thoát bào (exocytosis). Các protein khác cần gắn vào Ig như các chuỗi J cũng đồng thời được thực hiện trước khi bài tiết ra khỏi tế bào. Trong những tế bào sản xuất kháng thể lớp IgA và IgM thì sự sao chép của gen chuỗi nặng và gen chuỗi nhẹ được thực hiện đồng thời với sự sao chép gen chuỗi J.
Sự trưởng thành của các tế bào B trong tủy xương thường đi kèm với sự thay đổi biểu hiện gen Ig để dẫn đến sản xuất các phân tử Ig dưới các dạng khác nhau (Hình 3.2). Các tế bào sớm nhất của dòng lymphô B được gọi là tế bào tiền B sản xuất ra các dạng màng của chuỗi nặng μ, nhưng đa số các protein ở lại trong bào tương. Hầu hết các chuỗi nặng μ nội bào trong tế bào tiền B đều tự phân hủy ngay trong tế bào. Chỉ một lượng nhỏ các phân tử này được biểu hiện ở màng tế bào và kết hợp với những protein có tên là chuỗi nhẹ thay thế để tạo nên thụ thể của tế bào tiền B. Các tế bào B træåíng thành hoặc chưa trưởng thành đều sản xuất các chuỗi nhẹ κ hoặc λ để liên kết với chuỗi nặng μ tạo nên phân tử IgM. Sự tổ hợp này bảo vệ cho chuỗi nặng khỏi bị phân hủy trong nội bào và cho phép IgM biểu hiện trên bề mặt tế bào. Tế bào B trưởng thành có biểu hiện của IgM và IgD trên bề mặt. Các thụ thể Ig màng này có chức năng nhận diện kháng nguyên và khởi động quá trình hoạt hóa tế bào B. Thụ thể kháng nguyên của tế bào B liên kết không đồng hóa trị với hai protein màng khác là Igα và Igβ để tạo nên các phức hợp phân tử và điều này rất cần đối với việc thể hiện IgM và IgD bề mặt.
Khi tế bào B được kháng nguyên hoặc các kích thích khác hoạt hóa, chúng biệt hóa thành các tế bào sản xuất kháng thể. Quá trình này thường phối hợp với sự biến đổi các thể dạng sản xuất Ig. Một trong những biến đổi đó là sự chuyển từ Ig màng sang Ig tiết. Một biến đổi thứ hai là xuất hiện các biểu hiện khác của isotyp chuỗi nặng ngoài IgM và IgD. Quá trình này gọi là chuyển mạch isotyp chuỗi nặng.
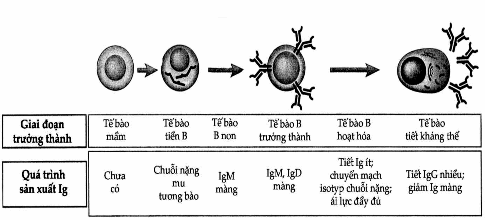
Hình. Sự bộc lộ Ig trong quá trình trưởng thành tế bào B.
Các giai đoạn trưởng thành tế bào B thường đi kèm với các biến đổi trong sản xuất chuỗi nặng và chuỗi nhẹ của Ig.
Bài viết cùng chuyên mục
Kỹ thuật DNA tái tổ hợp và miễn dịch lâm sàng
Kỹ thuật DNA tái tổ hợp, dựa vào việc sử dụng plasmid như các phương tiện để truyền các đoạn DNA lạ, ví dụ gen người.
Sản xuất kháng huyết thanh cho các phòng thí nghiệm miễn dịch lâm sàng
Hỗn dịch tế bào lách của con vật được gây mẫn cảm, có chứa nhiều tế bào B chịu trách nhiệm sản xuất nhiều kháng thể, chống nhiều epitope khác nhau.
Định typ HLA miễn dịch
Hiện nay, việc định týp HLA được thực hiện bằng kỹ thuật PCR, để phát hiện gen HLA, kỹ thuật này tốn kém hơn nhưng có độ chính xác và độ nhạy cao hơn.
Đánh giá tế bào trung tính và tế bào mono
Thực bào là chức năng ăn vật lạ của một tế bào nào đó, khả năng ăn này có thể xác định được bằng cách ủ tế bào thực bào, với các hạt trơ như hạt latex, hoặc vi khuẩn.
Khảo sát lymphô bào miễn dịch
Có hai loại phản ứng da in vivo, được dùng để phát hiện lympho bào T mẫn cảm đặc hiệu, đó là: thử nghiệm nội bì, dùng kháng nguyên tiêm vào lớp nội bì.
Khảo sát phức hợp miễn dịch
Hiện nay, trên thế giới, ta đã có bán những sản phẩm chuẩn, cho những phòng thí nghiệm miễn dịch đặc biệt chuyên khoa.
Khảo sát bổ thể miễn dịch
Định lượng C3 và C4, bằng phương pháp hóa miễn dịch, là các xét nghiệm có ích nhất, trên thế giới hiện nay đã sẵn có các huyết thanh chứng quốc tế.
Phát hiện tự kháng thể
Yếu tố thấp một kháng thể IgM, phản ứng với Ig,G đóng vai trò kháng nguyên sẽ cho phản ứng với IgG, ngưng kết mạnh hơn là với IgG người còn nguyên bản.
Kháng thể đối với kháng nguyên ngoại sinh
Thử nghiệm kích thích provocation test, tức thử nghiệm kích thích niêm mạc mũi hoặc niêm mạc phế quản bằng kháng nguyên, là một thử nghiệm khá phổ biến.
Khảo sát định tính immunoglobulin
Trong trường hợp không có bất thường chuỗi nặng, kháng huyết thanh chuỗi nhẹ tự do, tức không phản ứng với chuỗi nhẹ cố định vào chuỗi nặng.
Định lượng immunoglobulin và các protein đặc hiệu khác
Kỹ thuật thường được dùng phổ biến nhất, là miễn dịch kết tủa, tủa miễn dịch được hình thành khi kháng nguyên, và kháng thể, kết tủa tương ứng.
Đại cương các kỹ thuật miễn dịch
Một số xét nghiệm sẽ trở nên vô ích, nếu chúng ta yêu cầu không đúng lúc, đúng chỗ, các phân chia sẽ giúp lâm sàng có được chỉ định thích hợp.
Một số hiệu quả thuận lợi của phản ứng tự miễn
Tìm được những chất tải, hiệu quả nhất cũng như các tá chất thích hợp, để có thể gây phản ứng ở mức độ mong muốn.
Điều trị bệnh miễn dịch
Phản ứng tự miễn cũng có thể xảy ra nếu tế bào T hoặc B tự phản ứng mang một idiotyp phổ biến có phản ứng chéo với idiotyp trên kháng thể hoặc với cấu trúc trên vi khuẩn.
Chẩn đoán và tiên lượng bệnh miễn dịch
Tự kháng thể cón có giá trị tiên lượng, trường hợp một đứa trẻ, có anh chị em mắc bệnh đái đường phụ thuộc insulin, nó có chung HLA với anh chị.
Cơ chế bệnh sinh bệnh miễn dịch
Tế bào B tự phản ứng, tế bào T hiệu quả và tự kháng nguyên bình thường, vẫn có mặt trong cơ thể, nhưng không được khởi động.
Bệnh nguyên bệnh miễn dịch
Có những mô hình bệnh tự miễn ngẫu nhiên, trên động vật, rất có ích cho chúng ta nghiên cứu, đầu tiên là bệnh tuyến giáp tự miễn trên gà trống Obese.
Tính di truyền của bệnh tự miễn
Hoạt động của yếu tố di tryền, là xu hướng phối hợp của bệnh tự miễn, đối với các tính đặc hiệu HLA, Haplotyp B8, DR3 xuất hiện với tần suất cao.
Sự sắp xếp các bệnh tự miễn thường gặp ở người
Những cơ quan đích thường gặp, trong bệnh đặc hiệu cơ quan, là tuyến giáp, tuyến thượng thận, dạ dày và tuyến tụy.
Bằng chứng về tính chất gây bệnh của phản ứng tự miễn
Khi có được dòng động vật cảm thụ thích hợp rồi, ta còn có thể gây bệnh thụ động bằng cách truyền tế bào T mẫn cảm cho con vật khác.
Tính tự miễn dịch
Sự hình thành một đáp ứng kháng thể bình thường, đối với đa số kháng nguyên protein đòi hỏi sự tham gia của 3 loại tế bào B, T, và tế bào trình diện kháng nguyên.
Thiếu hụt miễn dịch thứ phát
Các thuốc ức chế miễn dịch, tác động lên rất nhiều khâu của chức năng tế bào, chức năng của lymphô bào và bạch cầu múi thường giảm.
Thiếu hụt miễn dịch tiên phát
Đối với những trường hợp thiếu hụt kháng thể bẩm sinh, nhiễm trùng tái đi tái lại bắt đầu xuất hiện, trong khoảng thời gian từ tháng thứ 4 đến 2 tuổi.
Đại cương thiếu hụt miễn dịch
Một đứa trẻ, hay một người lớn, sẽ bị nghi ngờ là thiếu hụt miễn dịch, khi trên cơ thể xuất hiện những nhiễm trùng lặp đi lặp lại, tồn tại kéo dài, trầm trọng hoặc bất thường.
Các phản ứng quá mẫn không đặc hiệu
Một cơ chế không đặc hiệu khác đã tham gia gây quá mẫn, đó là trường hợp thiếu các protein bất hoạt C3b, làm cho phản ứng hoạt hóa bổ thể không dừng lại.
Quá mẫn miễn dịch typ V (Quá mẫn kích thích)
Trên thực nghiệm, người ta đã phát hiện được thêm một số kháng thể kích thích hoạt động chức năng của tế bào, qua các kháng nguyên có trên bề mặt tế bào.
Quá mẫn miễn dịch typ IV (Quá mẫn muộn)
Cần luôn nhớ rằng, tổn thương quá mẫn, là hậu quả của phản ứng quá mức, giữa kháng nguyên, với cơ chế miễn dịch tế bào vẫn còn bình thường
Quá mẫn miễn dịch typ III
Kháng thể và kháng nguyên tạo thành phức hợp, phức hợp này hoạt hóa bổ thể đồng thời tác động gây giải phóng các amin hoạt mạch, làm tăng tính thấm thành mạch.
Quá mẫn miễn dịch typ II
Phản ứng truyền máu đối với các thành phần khác của máu, như bạch cầu, tiểu cầu cũng có thể xảy ra, nhưng hậu quả của nó không nặng nề như phản ứng đối với hồng cầu.
Quá mẫn miễn dịch typ I
Một khi IgE gắn thụ thể Fcε trên tế bào mast, và tế bào ái kiềm, sự mất hạt sẽ xảy ra khi có liên kết chéo, giữa các phân tử IgE.
Đại cương về quá mẫn miễn dịch
Qúa mẫn là một đặc điểm của cá thể, và nó xảy ra khi có tiếp xúc với kháng nguyên lần thứ hai.
Miễn dịch chống nấm
Nhiễm nấm có thể gây nhiều hậu quả, thường chỉ đáp ứng miễn dịch đặc hiệu chống nấm cùng với thuốc chống nấm tại chỗ, có thể tiêu diệt được bệnh nhiễm nấm cạn.
Miễn dịch chống ký sinh trùng
Bệnh nhân chống chọi với nhiễm ký sinh trùng đơn bào, bằng những phản ứng tương tự như trong nhiễm vi khuẩn, có một số đơn bào có cơ chế tồn tại độc đáo.
Miễn dịch chống vi khuẩn
Một số vi khuẩn xâm nhập qua đường niêm mạc, có thể tạo ra các protease để ly giải kháng thể IgA tiết, neisseria gonorrhea, neisseria meningitis.
Miễn dịch chống virus
Virus Epsptein-Barr dùng thụ thể của C3b tức CR2, còn HIV thì lại dùng thụ thể CD4 để làm nơi xâm nhập vaof tế bào đích, trong hệ thống miễn dịch.
Đại cương miễn dịch chống vi sinh vật
Đối với nhiễm trùng, một cân bằng được duy trì giữa sức chống đỡ của cơ thể, và khả năng của vi sinh vật cố gắng để vượt qua sức chống đỡ đó.
Cytokin kích thích tạo máu
Các cytokin khác nhau kích thích, sự phát triển, và trưởng thành của nhiều dòng tế bào máu khác nhau.
Các cytokin trung gian và điều hòa miễn dịch thu được
Mặc dù lúc đầu người ta phát hiện ra IL 2, như là một yếu tố phát triển tế bào T, nhưng thật ra IL 2 có nhiều chức năng trong đáp ứng miễn dịch thu được.
Các cytokin trung gian và điều hòa miễn dịch bẩm sinh
Có 2 loại thụ thể của TNF, loại có trọng lượng phân tử 55 kD có tên là THF RI, và loại có trọng lượng phân tử 75 kD có tên là TNF RII.
Đại cương về Cytokin
Các cytokin này do các tế bào đệm, bạch cầu, và một vài tế bào khác của tủy xương sản xuất, chúng có thể kích thích sự phát triển, và biệt hóa của bạch cầu non.
Sự hình thành phức hợp tấn công màng C5 9
Sau khi C5b gắn màng, C6 và C7 đến gắn vào C5b để tạo C5b67, C5b67 tác động với C8 để tạo C5b678, đơn vị này tạo phản ứng trùng hợp phân tử C9.
Con đường hoạt hóa bổ thể không cổ điển (alternative pathway)
Không nhất thiết phải có phản ứng kháng nguyên, kháng thể, nên phản ứng có thể xảy ra tức thì, và cơ thể được bảo vệ theo cơ chế không đặc hiệu.
Con đường hoạt hóa bổ thể cổ điển (classical pathway)
Một phân tử IgM pentamer, kết hợp với kháng nguyên, là có thể cố định bổ thể, nhưng đối với IgG, thì phải có phân tử IgG được gắn với kháng nguyên ở vị trí gần nhau.
Đại cương bổ thể
Các protein của hệ thống bổ thể tạo thành hai chuỗi enzym, mà người ta gọi là con đường cổ điển, và con đường không cổ điển, để tạo nên hai cách phân cách hai phân tử C3
Tế bào trình diện kháng nguyên
Tế bào hình sao được tìm thấy dưới lớp biểu bì, và trong đa số các cơ quan, ở đó chúng được đặt ở tư thế sẵn sàng để bắt giữ kháng nguyên.
Tế bào lymphô hệ miễn dịch
Tế bào lymphô bao gồm nhiều tiểu quần thể khác nhau, khác biệt về chức năng, về sản phẩm protein nhưng không phân biệt được về hình thái.
Đại cương các tế bào chủ yếu của hệ thống miễn dịch
Tế bào tham gia vào đáp ứng miễn dịch thu được, bao gồm các lymphô bào đặc hiệu kháng nguyên, tế bào trình diện kháng nguyên.
Một số kháng nguyên quan trọng
Người ta biết nhiều về cấu trúc của vùng H 2I của chuột, hơn vùng tương đương, với vùng này ở người là vùng HLA D.
Các tính chất của kháng nguyên
Một kháng nguyên protein phức tạp, có thể nhiều quyết định kháng nguyên khác nhau, do đó mà nó có thể kích thích tạo ra nhiều loại kháng thể khác nhau.
Sự kết hợp kháng nguyên kháng thể
Khả năng trung hòa độc tố, và vi sinh vật của kháng thể, luôn phụ thuộc vào sự gắn kết chặt chẽ của chúng vào kháng thể.
Phân bố tự nhiên và sản xuất kháng thể
Một người lớn khoẻ mạnh nặng 70 kg sản xuất khoảng 3g kháng thể mỗi ngày, khoảng 2 phần 3 là kháng thể có tên là IgA, được sản xuất bởi tế bào B.
Các con đường và cơ chế tái tuần hoàn và homing của tế bào lympho
Tế bào T nguyên vẹn có xu hướng homing, và tái tuần hoàn qua các cơ quan lymphô ngoại biên, để ở đó chúng sẽ nhận dạng, và đáp ứng với kháng nguyên.
Các cơ quan mô lymphô của hệ thống miễn dịch
Cơ quan và mô lymphô ngoại biên, bao gồm hạch bạch huyết, lách, hệ thống miễn dịch da, và hệ thống miễn dịch niêm mạc.
Các giai đoạn của đáp ứng miễn dịch thu được
Những nguyên lý cơ bản của thuyết chọn clôn, đã dần được chứng minh một cách thuyết phục, qua nhiều thí nghiệm, và tạo nên nền tảng cho quan niệm hiện nay.
Các thành phần tế bào của hệ thống miễn dịch thu được
Có một nhóm tế bào lymphô thứ ba, là tế bào giết, đây là những tế bào tham gia vào hệ thống miễn dịch bẩm sinh, chống lại nhiễm trùng virus.
Các đặc điểm chính của đáp ứng miễn dịch thu được
Tất cả đáp ứng miễn dịch bình thường, sẽ phai nhạt dần theo thời gian, để trả lại hệ miễn dịch ở trạng thái nghỉ ban đầu, tình trạng này gọi là hằng định nội môi.
Các kiểu đáp ứng miễn dịch thu được
Sự nhấn mạnh về kháng thể, trong lý thuyết này, đã dẫn đến sự thừa nhận chung, tầm quan trọng của miễn dịch dịch thể.
Tính miễn dịch bẩm sinh và thu được
Các cơ chế của miễn dịch tự nhiên, cung cấp sức đề kháng ban đầu đối với nhiễm trùng, đáp ứng miễn dịch thu được đến muộn hơn, với sự hoạt hoá tế bào lymphô.
Đại cương miễn dịch
Chức năng sinh lý của hệ thống miễn dịch, là bảo vệ một cơ thể chống lại các vi sinh vật gây bệnh, xâm nhập vào cơ thể đó.