- Trang chủ
- Sách y học
- Bài giảng miễn dịch
- Con đường hoạt hóa bổ thể cổ điển (classical pathway)
Con đường hoạt hóa bổ thể cổ điển (classical pathway)
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Quá trình enzym và tấn công màng dẫn đến giết chết tế bào. Đơn vị nhận diện bổ thể hoạt động của con đường này có thể chia làm ba giai đoạn: nhận dạng, hoạt hóa là phức hợp C1.
Cố định C1 bởi immunoglobulin
Cố định C1 xảy ra khi tiểu đơn vị C1q gắn trực tiếp lên Ig. Hai tiểu đơn vị còn lại của C1 là Clr và Cls không kiên kết với Ig mà tham gia vào quá trình hoạt hóa cổ điển về sau. Phân tử kháng thể sau khi kết hợp với kháng nguyên tương ứng sẽ bị thay đổi cấu trúc không gian làm lộ các thụ thể đối với C1q. Tính chất kết hợp của bổ thể vào phức hợp kháng nguyên - kháng thể là cơ sở của phản ứng cố định bổ thể mà chúng ta thường hay sử dụng trong các xét nghiệm miễn dịch học.
Không phải tất cả các Ig có khả năng kết hợp với bổ thể. Ở người chỉ có IgG1, IgG3, và IgM là có khả năng này; ở chuột nhắt có IGg2 và IgM, và chuột lang có IgG2, .v.v. Cấu hình không gian của phức hợp kháng nguyên - kháng thể cũng có ảnh hưởng đến khả năng cố định bổ thể. Nhưng cụ thể thì chưa được biết rõ.
Một phân tử IgM pentamer kết hợp với kháng nguyên là có thể cố định bổ thể nhưng đối với IgG thì phải có phân tử IgG được gắn với kháng nguyên ở vị trí gần nhau mới có thể cố định được bổ thể. Để đạt được điều này trong thực nghiệm đối với hồng cầu cừu, người ta tính ra phải cần đến 600-800 phân tử IgG gắn lên hồng cầu. Do đó mà người ta thường dùng IgM hơn. Vị trí gắn của bổ thể vào Ig là lĩnh vực CH2 của IgG và CH4 của IgM. Sau khi C1q đã kết hợp với kháng thể, sự hoạt hóa sẽ đựơc truyền sang C1r và Cls. Cơ chế C1q hoạt hóa C1r vẫn còn chưa được biết. C1s sau khi được hoạt hóa C4 và C2 là hai thành phần tiếp theo của chuỗi phản ứng.
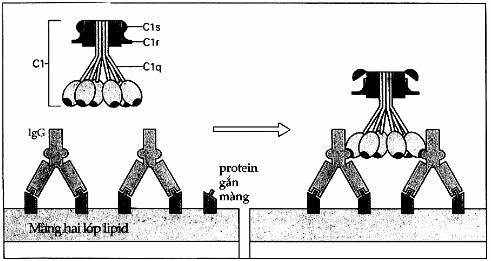
Hình. Cố định C1qrs.
Một cặp phân tử IgG liên kết với các phân tử protein kháng nguyên (protein gắn màng). Mảnh C1 cố định các lĩnh vực CH2 của IgG. Sự hoạt hóa C1r và C1s là do phân cắt nội bộ và do sự biến đổi của thành phần C1q tạo nên.
Cố định và hoạt hóa C4 và C2 bởi phức hợp C1qrs
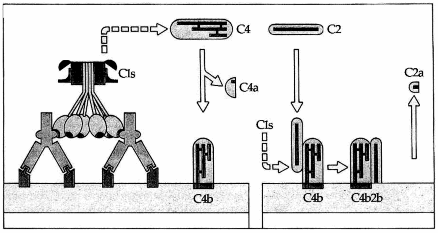
Hình. Sự hình thành C4b2b.
C1s cắt C4 tạo ra C4a và C4b. C4b gắn vào màng tế bào. C2 huyết thanh đến gắn vào C4b và bị C1s cắt để giải phóng C2a. C4b2b chính là enzym cắt C3 của con đường hoạt hóa cổ điển.
Phân tử C1s sau khi được hoạt hóa nó trở thành một enzym và cắt phân tử C4 thành hai mảnh: mảnh nhỏ là C4a và mảnh lớn C4b. Mảnh C4b có một vị trí hoạt động giúp nó gắn lên thụ thể dành cho nó trên bề mặt một số tế bào. Chúng ta cần lưu ý rằng mảnh C4b không nhất thiết chỉ nằm chung quanh nơi nó được hình thành mà nó thể vào máu tuần hoàn và được lưu động, tuy nhiên khi nó vào tuần hoàn thì trở nên bị bất hoạt. Phần C4b còn lại tại chäù (khoảng 10%) sẽ gắn lên tế bào.
C1s có tác dụng rất yếu lên C2 tự do, nhưng khi C2 tạo phức hợp với C4b thì tác dụng này mạnh hơn nhiều. Sự hấp thụ C2 lên C4b chỉ xảy ra trong điều kiện có mặt ion Mg++. C2 sẽ bị cắt làm hai mảnh, mảnh nhỏ C2a sẽ bị mất vào môi trường chung quanh còn mảnh lớn C2b thì vẫn còn bám trên C4b tạo thành phức hợp C4b2b có tính enzym không bền vững, có thời gian nửa đời sống ngắn (5 phút) vì mảnh C2b dễ bị hủy hoại hoặc tách rời khỏi C4b (Hình 6.2).
Tác động của C4b2b lên C3
Phức hợp C4b2b hoạt hóa phân tử C3 bằng cách cắt phân tử này thành hai mảnh C3a và C3b. Mảnh C3a có trọng lượng phân tử thấp, được giải phóng vào trong dịch cơ thể và có hoạt tính của một hóa chất trung gian của phản viêm. Mảnh lớn của C3b có trọng lượng phân tử khoảng 175.000 kD và có khả năng dính lên bề mặt tế bào. Tuy nhiên, chỉ có tỷ lệ nhỏ C3b dính được vào màng tế bào ở vị trí sát với phức hợp enzym C4b2b để tạo phức hợp C4b2b3b có diện tác dụng nằm trên mảnh C3b. Các phân tử C3b bám vào màng tế bào ở các vị trí xa C4b2b thì chỉ có vai trò giúp cho việc kết dính miễn dịch và làm dễ hiện tượng thực bào nhờ vào đầu kia gắn lên thụ thể dành cho C3 ở các thực bào.
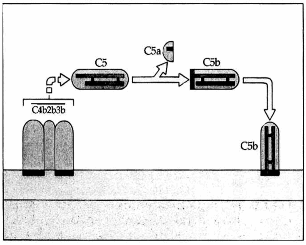
Hình. Cố định C5.
C4b2b3b tách mảnh C5a từ chuỗi α của C5 ra. Mảnh còn lại là C5b sau đó gắn vào màng tế vào.
Tác động của C3b lên C5
Phức hợp C4b2b3b mà diện tác dụng là C3b cắt phân tử C5 thành ra hai mảnh. Mảnh C5a là mảnh nhỏ cũng được giải phóng vào dịch cơ thể và có tác dụng như một chất trung gian hóa học của phản ứng viêm. Mảnh C5b sẽ được cố định lên màng tế bào và bắt đầu tạo phức hợp tấn công màng.
Bài viết cùng chuyên mục
Miễn dịch chống nấm
Nhiễm nấm có thể gây nhiều hậu quả, thường chỉ đáp ứng miễn dịch đặc hiệu chống nấm cùng với thuốc chống nấm tại chỗ, có thể tiêu diệt được bệnh nhiễm nấm cạn.
Đại cương các kỹ thuật miễn dịch
Một số xét nghiệm sẽ trở nên vô ích, nếu chúng ta yêu cầu không đúng lúc, đúng chỗ, các phân chia sẽ giúp lâm sàng có được chỉ định thích hợp.
Một số hiệu quả thuận lợi của phản ứng tự miễn
Tìm được những chất tải, hiệu quả nhất cũng như các tá chất thích hợp, để có thể gây phản ứng ở mức độ mong muốn.
Đại cương miễn dịch
Chức năng sinh lý của hệ thống miễn dịch, là bảo vệ một cơ thể chống lại các vi sinh vật gây bệnh, xâm nhập vào cơ thể đó.
Quá mẫn miễn dịch typ II
Phản ứng truyền máu đối với các thành phần khác của máu, như bạch cầu, tiểu cầu cũng có thể xảy ra, nhưng hậu quả của nó không nặng nề như phản ứng đối với hồng cầu.
Sản xuất kháng huyết thanh cho các phòng thí nghiệm miễn dịch lâm sàng
Hỗn dịch tế bào lách của con vật được gây mẫn cảm, có chứa nhiều tế bào B chịu trách nhiệm sản xuất nhiều kháng thể, chống nhiều epitope khác nhau.
Tính di truyền của bệnh tự miễn
Hoạt động của yếu tố di tryền, là xu hướng phối hợp của bệnh tự miễn, đối với các tính đặc hiệu HLA, Haplotyp B8, DR3 xuất hiện với tần suất cao.
Các cơ quan mô lymphô của hệ thống miễn dịch
Cơ quan và mô lymphô ngoại biên, bao gồm hạch bạch huyết, lách, hệ thống miễn dịch da, và hệ thống miễn dịch niêm mạc.
Định lượng immunoglobulin và các protein đặc hiệu khác
Kỹ thuật thường được dùng phổ biến nhất, là miễn dịch kết tủa, tủa miễn dịch được hình thành khi kháng nguyên, và kháng thể, kết tủa tương ứng.
Miễn dịch chống ký sinh trùng
Bệnh nhân chống chọi với nhiễm ký sinh trùng đơn bào, bằng những phản ứng tương tự như trong nhiễm vi khuẩn, có một số đơn bào có cơ chế tồn tại độc đáo.
Miễn dịch chống vi khuẩn
Một số vi khuẩn xâm nhập qua đường niêm mạc, có thể tạo ra các protease để ly giải kháng thể IgA tiết, neisseria gonorrhea, neisseria meningitis.
Một số kháng nguyên quan trọng
Người ta biết nhiều về cấu trúc của vùng H 2I của chuột, hơn vùng tương đương, với vùng này ở người là vùng HLA D.
Các giai đoạn của đáp ứng miễn dịch thu được
Những nguyên lý cơ bản của thuyết chọn clôn, đã dần được chứng minh một cách thuyết phục, qua nhiều thí nghiệm, và tạo nên nền tảng cho quan niệm hiện nay.
Cấu trúc phân tử của kháng thể
Các glycoprotein huyết tương, hoặc huyết thanh trước đây thường được phân chia theo tính chất hòa tan, của chúng thành albumin và globulin.
Các thành phần tế bào của hệ thống miễn dịch thu được
Có một nhóm tế bào lymphô thứ ba, là tế bào giết, đây là những tế bào tham gia vào hệ thống miễn dịch bẩm sinh, chống lại nhiễm trùng virus.
Đại cương các tế bào chủ yếu của hệ thống miễn dịch
Tế bào tham gia vào đáp ứng miễn dịch thu được, bao gồm các lymphô bào đặc hiệu kháng nguyên, tế bào trình diện kháng nguyên.
Đại cương về Cytokin
Các cytokin này do các tế bào đệm, bạch cầu, và một vài tế bào khác của tủy xương sản xuất, chúng có thể kích thích sự phát triển, và biệt hóa của bạch cầu non.
Quá mẫn miễn dịch typ I
Một khi IgE gắn thụ thể Fcε trên tế bào mast, và tế bào ái kiềm, sự mất hạt sẽ xảy ra khi có liên kết chéo, giữa các phân tử IgE.
Chẩn đoán và tiên lượng bệnh miễn dịch
Tự kháng thể cón có giá trị tiên lượng, trường hợp một đứa trẻ, có anh chị em mắc bệnh đái đường phụ thuộc insulin, nó có chung HLA với anh chị.
Miễn dịch chống virus
Virus Epsptein-Barr dùng thụ thể của C3b tức CR2, còn HIV thì lại dùng thụ thể CD4 để làm nơi xâm nhập vaof tế bào đích, trong hệ thống miễn dịch.
Thiếu hụt miễn dịch thứ phát
Các thuốc ức chế miễn dịch, tác động lên rất nhiều khâu của chức năng tế bào, chức năng của lymphô bào và bạch cầu múi thường giảm.
Tính miễn dịch bẩm sinh và thu được
Các cơ chế của miễn dịch tự nhiên, cung cấp sức đề kháng ban đầu đối với nhiễm trùng, đáp ứng miễn dịch thu được đến muộn hơn, với sự hoạt hoá tế bào lymphô.
Sự kết hợp kháng nguyên kháng thể
Khả năng trung hòa độc tố, và vi sinh vật của kháng thể, luôn phụ thuộc vào sự gắn kết chặt chẽ của chúng vào kháng thể.
Khảo sát bổ thể miễn dịch
Định lượng C3 và C4, bằng phương pháp hóa miễn dịch, là các xét nghiệm có ích nhất, trên thế giới hiện nay đã sẵn có các huyết thanh chứng quốc tế.
Cơ chế bệnh sinh bệnh miễn dịch
Tế bào B tự phản ứng, tế bào T hiệu quả và tự kháng nguyên bình thường, vẫn có mặt trong cơ thể, nhưng không được khởi động.
