- Trang chủ
- Sách y học
- Bài giảng sinh lý bệnh
- Chu kỳ tế bào và cơ chế sửa sai trong sao chép
Chu kỳ tế bào và cơ chế sửa sai trong sao chép
Bộ gen tuyến trùng Caenorhabditis elegans có 1031 gen, trong đó có 131 gen có phân định sẳn là chết theo chương trình.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Chu kỳ tế bào
Một chu kỳ sinh học tế bào tức là giai đoạn giữa hai lần phân chia gồm 4 pha: M, G1, S, G2. Chu kỳ phân chia kéo dài khoảng 16-24 giờ tuỳ theo mỗi loại tế bào.
M (mitosis): Hoạt động phân chia tế bào hay nhân đôi tế bào
G1 (gap): Có sự tích luỹ vật chất nội bào và năng lượng, kết thúc ở điểm tới hạn R (restriction) vài giờ trước khi chuyển từ G1 sang S. Một khi tế bào đi qua được điểm R sẽ đi qua các pha khác để thực hiện được phân bào.
S (synthesis): Giai đoạn tổng hợp DNA, lượng DNA tăng gấp đôi (từ 23 đôi thành 46 đôi)
G2: Quy trình được hoàn tất và chuẩn bị sang pha sau
M (mitosis): Mỗi cặp kép nhiễm sắc thể chia đôi, đi về 2 cực tạo thành 2 tế bào con y hệt tế bào mẹ.
Sau khi phân đôi 2 tế bào con có thể tiếp tục chu trình ấy hoặc đi vào thời kỳ nghỉ là G0.
G0: Các thời gian dừng của chu kỳ tế bào là để sửa chữa DNA cho tế bào sống sót và không tiến triển thành ung thư: sự ngừng chu kỳ tế bào ở G1/S tránh được sự tái sao của các DNA thương tổn, sự dừng ở G2/M tránh được sự ngưng tập của các nhiễm sắc thể bị thương tổn.
Cơ chế sửa sai trong chu kỳ tế bào
Ngay từ những năm 1960 Leland Hartwell đã phân lập được nhiều loại tế bào có đột biến gen ở một loại nấm men (Saccharomyces cerevisae). Bằng cách tái hợp chúng với nhau đã phát hiện ra sự kiểm soát phân chia tế bào nằm trên NST do hằng trăm gen khác nhau. tên gọi chung là gen CDC (cell division cycle genes) với chữ số theo sau cho từng thứ. Trong số các gen bày có hai loại quan trọng nhất là điểm khởi phát chu kỳ và những điểm quan trọng mà khi bị hư hỏng thì gián đoạn phân chia tế bào, gọi là điểm kiểm soát (check point)
Sửa sai trong sao chép
Thực nghiệm dùng các nucleotid, DNA polymerase để tổng hợp DNA thì nhận thấy sai sót xảy ra rất cao (1x10-5) trong khi sao chép tự nhiên lại thấp hơn nhiều. DNA của E. coli có 3x106 cặp bazơ như vậy thì mỗi lần sao chép phải có 30 sai sót xảy ra dẫn đến sự đột biến nhưng điều này không xảy ra như vậy trong tự nhiên. Nguyên nhân chính của sự chính xác này là hiện tượng sửa chữa DNA xảy ra ở mọi tế bào bình thường trong cơ thể sinh vật bậc cao.
Theo dõi tần số đột biến ở các quần thể lớn cho thấy tỷ lệ đột biến chỉ ở 1x10-9, như vậy ở người mỗi lần sao chép chỉ có 3 sai sót xảy ra cho mỗi DNA, như vậy kết luận cơ thể sinh vật đã có những cơ chế sửa sai tế bào.
Hàng chục loại enzym khác nhau đã tham gia vào quá trình sửa chữa các DNA tổn thương. Chúng nhận biết chọn lọc một bazơ bị thay đổi, loại bỏ nucleotid mang nó bằng cách cắt ra khỏi chuỗi DNA, sau đó thay bằng một nucleotid mang bazơ chính xác bổ sung và gắn DNA lại. Người ta cho rằng với cơ chế này cho phép sửa chữa 99,9% các sai sót.
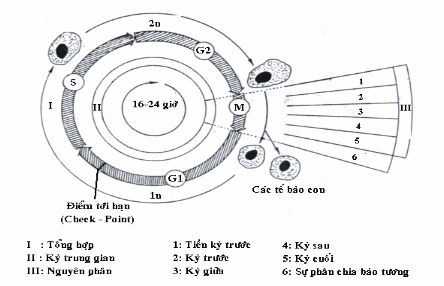
Hình: Chu kỳ tế bào.
Mở đầu khi tế bào chuyển sang pha G1, Harwell phát hiện là nó sẽ bị ngăn lại khi thiếu chất dinh dưỡng hoặc chất chuyển hoá trung gian, có sự tham gia của gen RNA polymerase (primase) tạo ra đoạn mồi đầu tiên để cho DNA polymerase tiếp tục quá trình tái sao DNA cũng như loại bỏ mẩu mồi ấy. Khi tế bào qua khỏi điểm này (điểm khởi phát, điểm R: restriction) thì sẽ không quay trở lại được và phải đi tiếp sang pha S. Những rối loạn xảy ra ở các bước sau tế bào sẽ bị loại bỏ bằng cơ chế chết theo chương trình (apoptosis).
Các sự kiện của chu kỳ tế bào diễn tiến theo một trật tự nhất định, sự kiện trước phải được hoàn tất tốt đẹp thì sự kiện sau mới tiếp tục xảy ra. Các cơ chế kiểm soát nhờ hoạt động của những gen nằm ở những nơi gọi là điểm kiểm soát. Khi các điểm kiểm soát bị loại bỏ sẽ gây chết tế bào, sai lệch trong phân bố nhiễm sắc thể hay các phần tử tế bào hoặc tăng nhạy cảm với các yếu tố môi trường.
Sau này Harwell đã tìm ra được nhiều gen của điểm kiểm soát. Ví dụ như tế bào nấm có gen RAD53 khi bị chiếu tia thì sẽ gây ngừng ở G2 cho đến khi tổn thương được sửa chữa xong. Ở người có gen p53 trên nhiễm sắc thể 17 có vai trò đièu hoà tương tự như RAD53. Khi gen này do đột biến không hoạt động được thì tế bào dù bị hư hại (đột biến hay thay đổi do yếu tố sinh ung thư) vẫn cứ thế mà tiếp tục phân chia thành những thế hệ tế bào con có rối loạn y hệt gọi là ác tính.
Năm 1970 nhờ các kỹ thuật gen và sinh học phân tử đã phát hiện ra khi vượt qua điểm khởi động thì tế bào cần hoạt động của 2 gen cdc2+/cdc28+ để bắt đầu tái sao DNA. Các gen này mã cho một protein kinase p34 là yếu tố điều hoà chủ yếu của phức hợp. Sau này người ta mới phát hiện ra vai trò của cyclin với p34cdc2 protein kinase
Năm 1980 R. Timothy Hunt đã phát hiện ra phân tử cyclin đầu tiên trong quá trình phân bào của con nhím biển, ức chế sự tổng hợp các protein này thì các giai đoạn phân bào không thực hiện được. Protein này hình thành rồi phân huỷ ngay trong mỗi chu kỳ tế bào nên mới có tên là cyclin (cycle). Hiện nay, cyclin tạo thành một họ gồm nhiều protein gần giống nhau có đặc tính chung là liên kết với các dưới đơn vị protein kinase trong họ cdc để trở nên các phức hợp hoạt động, gọi là protein kinase phụ thuộc cyclin (cdk). Để cho 1 pha chu kỳ tế bào đi qua được 1pha khác thì cdk này phải kết hợp với 1 cyclin nhất định.
Ví dụ:
Cyclin B + cdk1: giúp tế bào đi từ G2 sang M Cyclin E + cdk2: G1→S
Cyclin D + cdk4: G0 → G1
Các gãy đơn (single strand breaks) và gãy kép (double strand breaks) của DNA làm tăng tổng hợp p53, làm tế bào dừng lại ở pha G1do ức chế được phức hợp Cyclin E + cdk2 nên tế bào không đi vào pha S. Nếu thiếu p53, tế bào dễ phát triển thành ung thư.
Sửa sai khi không sao chép
Có khoảng 50 enzym chuyên rà soát, phát hiện và sửa các sai sót trên phân tử DNA.
Sự chết tế bào
Phân bào làm tăng số lượng tế bào với bộ gen giống nhau. Song song với sự phân chia tế bào còn có những cơ chế kiểm soát sự chết của tế bào theo chu kỳ phát triển. Có thể gặp trong một số quá trình sinh lý như thoái triển sinh lý phụ thuộc nội tiết tố, tế bào lympho T gây độc (Tc) làm chết tế bào đích....Ngoài ra, không kể đến những trường hợp chết tế bào do thoái hoá, hoại tử, sự teo ..v.v.
Chết theo chương trình của tế bào (apoptosis nghĩa đen là rơi khỏi)
Trên tiêu bản nhuộm hematoxylin eosin cho thấy có sự biến tính của protein, chromatin đậm đặc. Cuối cùng có hiện tượng vỡ nhân do DNA bị gãy, tế bào nhanh chóng co lại và tạo ra các chồi bào tương, đứt gãy thành các “thể apotosis” chứa chất hoà tan và cơ quan tử của tế bào; các thể này được thực bào sau đó. Cơ chế này vẫn chưa hoàn toàn rõ ràng tuy nhiên thường bắt đầu bằng các thương tổn DNA, đặc biệt là hiện tượng gãy DNA chuổi kép, tăng giải phóng topoisomerase (có trong tế bào bình thường hoặc ác tính) có tác dụng tách đôi sợi DNA xoắn kép khi sao chép, sửa chữa các thương tổn DNA. Khi thiếu enzym này thì nhiễm sắc thể không phân ly và tế bào không phân bào được. Nhưng khi quá nhiều topoisomerase thì tế bào cũng sẽ chết theo chương trình khi có sự kết hợp giữa DNA chuổi kép và topoisomerase II.
Tóm lại, ở sinh vật đa bào sự kiểm soát phân bào và chết theo chương trình được kiểm soát bằng nhiều cơ chế phức tạp. Khi cơ chế kiểm soát này bị hỏng thì dẫn đến nguyên phân không giới hạn và tế bào hầu như không biệt hoá, xảy ra ở ung thư.
Sự kiểm soát di truyền đối với apoptosis
Bộ gen tuyến trùng Caenorhabditis elegans có 1031 gen, trong đó có 131 gen có phân định sẳn là chết theo chương trình. Có 14 gen khác nhau tham gia vào kiểm soát di truyền đối với apotosis.
Nếu có sự đột biến làm bất hoạt các gen ức chế sinh ung thư (ced -3 và ced -4) thì các tế bào trên sống sót tuy nhiên nếu đột biến gen kích thích sinh ung thư (ced -9) làm cho một số tế bào bình thường sống sót nay bị chết. Nghiên cứu cấu trúc chức năng của một protein của gen ced-9 thì thấy giống như sản phẩm của một proto-oncogen có tên là bcl-2. Ngoài ra p53 là sản phẩm của gen ức chế ung thư, là chất hoạt hoá một trong những con đường dẫn đến apoptosis và làm giảm protein bcl-2.
Apoptosis và bệnh lý khác
Sự chết theo chương trình của tế bào giữ vai trò cơ bản trong nhiều bệnh: điều hoà trực tiếp sự phát triển của khối u, góp phần ngăn chận hay làm chậm đi sự phát triển của bệnh AIDS. Nhiều bằng chứng cho thấy vai trò của apoptosis trong tuổi thọ, bệnh Alzheimer,.vv. Tuy được phát hiện không lâu nhưng apoptosis đã thu hút những nghiên cứu về điều trị ung thư và những bệnh lý khác.
Ung thư
(xem bài ung thư).
Bài viết cùng chuyên mục
Cảm giác nhiệt: Receptor và sự hưng phấn của chúng
Những receptor nóng và lạnh nằm ngay dưới da ở những điểm tách biệt riêng rẽ. Hầu hết các vùng của cơ thể, các điểm lạnh gấp 3 đến 10 lần điểm nóng, và những vùng khác nhau thì có số điểm khác nhau.
Quá trình viêm: vai trò của bạch cầu hạt trung tính và dại thực bào
Viêm đặc trưng bởi sự giãn rộng của mạch tại chỗ, gây tăng lưu lượng máu tại chỗ; tăng tính thấm của mao mạch, cho phép rò rỉ một lượng lớn dịch vào khoảng kẽ; thường đông tụ dịch ở khoảng kẽ.
Hội chứng thận hư: mất protein theo nước tiểu và giữ natri
Do nồng độ protein huyết tương giảm, áp suất thẩm thấu keo huyết tương giảm xuống mức thấp. Điều này khiến các mao mạch trên khắp cơ thể lọc một lượng lớn dịch vào các mô khác nhau, do đó gây ra phù nề và giảm thể tích huyết tương.
Bất thường lâm sàng của điều chỉnh thể tích dịch: hạ và tăng natri máu
Hạ Na còn có thể do cơ thể hấp thu quá nhiều nước do tiết quá nhiều ADH, một hoocmon chống bài niệu, gây tái hấp thu quá nhiều nước vào cơ thể.
Nhắc lại sinh lý sinh hóa glucose máu
Tất cả các loại glucid đều được chuyển thành đường đơn trong ống tiêu hóa và được hấp thu theo thứ tự ưu tiên như sau: galactose, glucose, fructose và pentose.
Hệ thống đệm H+ trong dịch cơ thể
Tầm quan trọng của hệ thống đệm được thể hiện rõ khi nồng độ H+ thấp trong các dịch cơ thể và lượng tương đối lớn acid được sản xuất ra trong cơ thể.
Thận bài tiết nước dư thừa bằng cách hình thành nước tiểu loãng
Thận bình thường có một khả năng rất lớn để thay đổi tỷ lệ tương đối của các chất tan và nước trong nước tiểu để đáp ứng với những thách thức khác nhau.
Ống thận: sự bài thiết H+ và sự tái hấp thu HCO3-
Khoảng 80-90% HCO3- tái hấp thu (và H+ bài tiết) được thực hiện ở ống lượn gần, chỉ một số ít HCO3- xuống ống lượn xa và ống góp. Ở đoạn dày của nhánh lên quai Henle, khoảng 10% HCO3- nữa được tái hấp thu, và phần còn lại được hấp thu ở ống lượn xa và ống góp.
Đau đầu ngoài sọ: đau đầu do mắt và mũi
Sự căng thẳng cảm xúc thường làm co nhiều cơ của đầu, đặc biệt là cơ gắn vào da đầu và cơ cổ gắn với chẩm, trở nên co cứng, và nó được cho rằng cơ chế này là một trong những nguyên nhân thông thường của đau đầu.
Hệ thống đệm phosphat: điều chỉnh thăng bằng kiềm toan trong cơ thể
Hệ thống đệm phosphat có pK của 6.8, giá trị đó không xa pH bình thường trong dịch cơ thể là 7,4; điều này cho phép hệ thống đệm hoạt động gần tối đa.
Đa hồng cầu: tăng số lượng hồng cầu và hematocrit
Các nguyên bào không ngừng sản xuất hồng cầu khi đã đủ lượng hồng cầu. Điều này làm cho tăng quá mức sản xuất hồng cầu giống như trong ung thư vú sản xuất dư thừa của một loại tế bào vú nào đó.
Hồng cầu: sản sinh biệt hóa và tổng hợp
Hầu hết hồng cầu được sản xuất tại các xương có màng như xương cột sống, xương ức, các xương sườn và các xương chậu. Thậm chí, các xương này cũng giảm sản xuất khi tuổi tăng lên.
Kiểm soát dịch ngoại bào: các cơ chế của thận
Sự thay đổi lượng natri clorua trong dịch ngoại bào tương ứng với sự thay đổi tương tự lượng nước ngoại bào, và do đó duy trì nồng độ thẩm thấu và nồng độ natri tương đối ổn định.
Kích thích giải phóng ADH: do áp lực động mạch giảm và / hoặc thể tích máu giảm
Bất cứ khi nào huyết áp và lượng máu bị giảm, chẳng hạn như xảy ra trong xuất huyết, sự tăng tiết ADH dẫn đến tăng sự tái hấp thu dịch bởi thận, giúp khôi phục huyết áp và lượng máu về bình thường.
Đại cương sinh lý bệnh lão hóa
Tăng nhạy cảm với bệnh tật, tăng nguy cơ tử vong: hầu hết cơ thể già mang một hoặc nhiều bệnh và có tỷ lệ tử vong cao nhất so với mọi giai đoạn phát triển trước đó.
Độ chính xác của thể tích máu và điều chỉnh dịch ngoại bào
Sự thay đổi nhỏ trong huyết áp gây ra sự thay đổi lớn về lượng nước tiểu. Những yếu tố này kết hợp với nhau để cung cấp phản hồi kiểm soát lượng máu hiệu quả.
Một số quan niệm chưa đầy đủ về bệnh nguyên
Do không phân biệt được nguyên nhân và điều kiện hoặc không phân biệt được vai trò của mỗi yếu tố trong quá trình gây bệnh
Sinh lý bệnh của suy thượng thận và bệnh lý Addison
Lượng lớn của glucocorticoid đôi khi tiết ra để đáp ứng với các loại stress về thể chất hoặc tinh thần, bệnh Addison, đầu ra của glucocorticoid không tăng trong khi bị stress.
Sinh lý bệnh viêm cấp
Các tế bào và tiểu cầu thực hiện các chức năng với sự hỗ trợ của 3 hệ thống protein huyết tương đó là hệ thống bổ thể, hệ thống đông máu, hệ thống kinin.
Đối kháng thụ thể mineralocorticoid: giảm tái hấp thu natri và giản bài tiết kali của ống góp
Do các thuốc này cũng ức chế tác dụng của aldosterone trong việc làm tăng bài tiết kali, nên dẫn đến giảm bài tiết kali ra nước tiểu. Đối kháng thụ thể mineralocorticoid cũng làm kali từ các tế bào đi vào trong dịch ngoại bào.
Giải phẫu sinh lý của thận và hệ tiết niệu
Mặt giữa của mỗi quả thận chứa một vùng rốn, qua đó đi qua động mạch thận và tĩnh mạch, bạch huyết, cung cấp thần kinh và niệu quản, nơi mang nước tiểu cuối cùng từ thận đến bàng quang.
Đông máu nội mạch rải rác: tắc nghẽn mạch máu nhỏ ở ngoại vi
Sự tắc nghẽn các mạch máu nhỏ ngoại vi sẽ ngăn cản mô nhận oxy và các chất dinh dưỡng khác, điều này sẽ dẫn đến hoặc làm nặng thêm tình trạng shock tuần hoàn.
Nephron: đơn vị chức năng của thận
Mỗi nephron chứa một chùm mao mạch cầu thận được gọi là cầu thận, qua đó một lượng lớn dịch được lọc từ máu, và một ống dài trong đó dịch đã lọc được chuyển hóa thành nước tiểu.
Viêm nhiễm: sự đáp ứng của đại thực bào và bạch cầu hạt trung tính
Đại thực bào có thể thực bào nhiều vi khuẩn hơn (khoảng gấp 5 lần) và các phần tử lớn hơn, bao gồm cả bạch cầu hạt trung tính. Đại thực bào cũng đóng vai trò quan trọng trong việc khởi động sản xuất kháng thể.
Kiểm soát tuần hoàn thận của hormon và các chất hóa học
Trong trạng thái căng thẳng, chẳng hạn như sự suy giảm khối lượng hoặc sau khi phẫu thuật, các thuốc kháng viêm không steroid như aspirin, ức chế tổng hợp prostaglandin có thể gây ra giảm đáng kể mức lọc cầu thận.
