- Trang chủ
- Sách y học
- Sinh lý y học
- Tín hiệu nội bào sau khi receptor hormone được kích hoạt
Tín hiệu nội bào sau khi receptor hormone được kích hoạt
Nhiều loại mô khác nhau có cùng loại receptor nội bào giống nhau, tuy nhiên gen được kích thích bởi các receptor ở một số mô lại khác nhau.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Gần như không có ngoại lệ, một hormone tác dụng lên mô đích đầu tiên phải tạo ra phức hợp hormone-receptor. Dạng phức hợp này có chức năng khác với receptor, và sự kích hoạt receptor bắt đầu quá trình ảnh hưởng của hormone tại mô đích.
Receptor gắn trên kênh ion
Hầu hết tất cả các chất dẫn truyền thần kinh, như acetylcholine và nor-epinephrin, gắn vào các receptor màng sau synap. Sự liên kết này hầu như luôn luôn làm thay đổi cấu trúc của các receptor, thường mở hoặc đóng các kênh đối với một hoặc nhiều loại ion. Một số kênh ion phụ thuộc receptor mở (hoặc đóng) kênh với ion Natri, một số khác với ion Kali, một số khác thì với ion calci, và nhiều lạo khác nữa. Sự di chuyển khác nhau của các loại ion khác nhau vào và ra khỏi tế bào thông qua các kênh ion tạo ra những hiệu ứng kế tiếp trên tế bào sau synap. Mặc dù một vài hormone có thể thực hiện chức năng thông qua hoạt hóa receptor của các kênh ion, nhưng hầu hết các hormone mở hoặc đóng kênh ion gián tiếp thông qua việc gắn với receptor liên kết protein G hoặc các receptor liên kết enzyme.
Receptor liên kết protein G
Nhiều receptor sau khi được kích hoạt gián tiếp điều khiển hoạt động của các protein đích (như các enzyme hoặc các kênh ion) bằng cách gắn với nhóm các protein màng tế bào được gọi là heterotrimeric guanosine triphosphate (GTP)-gắn protein G. Đã phát hiên ra hơn 1000 receptor gắn protein G, tất cả đều có 7 phân đoạn xuyên màng lượng ra lượn vào xuyên qua màng tế bào. Một số phần của receptor gắn vào tế bào chất (đặc biệt là các đuôi tế bào chất của receptor) được liên kết với các protein G (protein G bao gồm 3 tiểu đơn vị: α, β và γ). Khi hormone gắn vào phần bên ngoài màng của receptor, sự thay đổi cấu hình của receptor làm kích hoạt protein G và phát ra các tín hiệu nội nào gây ra: hoặc (1) mở hoặc đóng các kênh ion màng, hoặc (2) thay đổi hoạt động của một enzyme trong tế bào chất, hoặc (3) kích hoạt phiên mã gen.
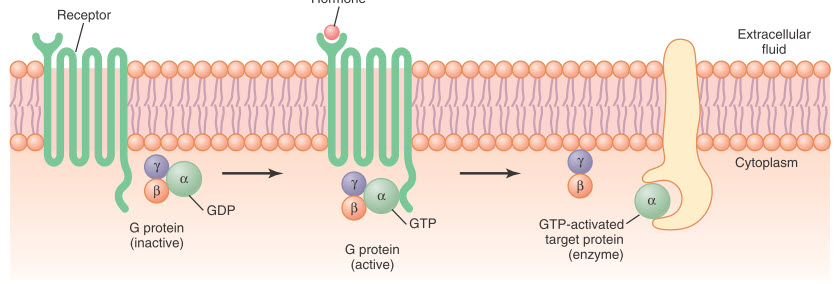
Hình. Cơ chế hoạt hóa thụ thể liên kết với protein G. Khi hormone kích hoạt thụ thể, các α, β và γ không hoạt động. Phức hợp protein G liên kết với thụ thể và được kích hoạt, với sự trao đổi guanosine triphosphate (GTP) cho guanosine diphosphate (GDP). Quá trình này làm cho tiểu đơn vị α (mà GTP được liên kết) phân ly khỏi tiểu đơn vị β và γ của protein G và tương tác với các protein đích gắn kết màng (enzym) bắt đầu các tín hiệu nội bào.
Protein tam phân G được đặt tên dựa trên khả năng gắn với guanosine nucleotides. Trong trạng thái bất hoạt của protein G, các tiểu đơn vị α, β và γ tạo thành một phức hợp gắn với guanosine diphosphate (GDP) tại vị trí tiểu đơn vị α. Khi receptor được kích hoạt, nó sẽ thay đổi hình dạng và làm cho phức hợp GDP-protein G liên kết với phần nằm trong tế bào chất của receptor và chuyển GDP thành GTP. Sự thay thế GDP thành GTP làm cho tiểu đơn vị α tách khỏi phức hợp tam phân và chuyển sang liên kết với một protein thông tin nội bào khác; những protein này, khi gắn với tiểu đơn vị α, sẽ làm thay đổi hoạt động của các kênh ion hoặc các enzyme nội bào như adenyl cyclase hoặc phospholipase C, làm biến đổi chức năng của tế bào.
Tín hiệu kết thúc khi hormone bị phá hủy và tiểu đơn vị α quay lại gắn với tiểu đơn vị β và γ để trở về dạng bất hoạt - chất điều tiết protein G.
Một số hormone được gắn với các protein G ức chế (kí hiệu là protein Gi), trong khi các hormone khác gắn với protein G kích thích (kí hiệu protein Gs). Do đó, tùy thuộc vào sự gắn của receptor hormone với protein G ức chê hay kích thích, một hormone có thể hoặc làm tăng hoặc giảm hoạt động của các enzyme nội bào. Phức hợp hệ thống protein G trên màng tế cung cấp một mảng rộng lớn của các tế bào tiềm năng đáp ứng lại các hormone khác nhau tại các mô khác nhau của cơ thể.
Receptor liên kết enzyme
Một số receptor, khi hoạt động mang chức năng giống như các enzyme hoặc liên kết chặt chẽ với các enzyme mà chúng kích hoạt. Những receptor liên kết enzyme này là những protein xuyên màng một lần, khác với các receptor xuyên màng gắn với protein G xuyên màng 7 lần. Receptor liên kết enzyme có những vị trí gắn hormone ở phía ngoài màng tế bào và tạo ra xúc tác hoặc vị trí gắn hormone ở bên trong. Khi hormone gắn vào phần ngoài màng của receptor, một enzyme trong màng tế bào ngay lập tức được kích hoạt (hoặc đôi khi bị bất hoạt). Mặc dù nhiều receptor liên kết enzyme có hoạt động của enzyme nội tại, một số khác phải cần đến các enzyme gắn chặt chẽ vào receptor mới có thể tạo ra sự thay đổi chức năng của tế bào.

Bảng Các hormone sử dụng tín hiệu Tyrosine Kinase của Receptor
Một số yếu tố tăng trưởng (có bản chất peptid), các cytokine và các hormone sử dụng receptor liên kết enzyme tyrosin kinase để tạo ra tín hiệu tế bào. Một ví dụ của receptor liên kết enzyme là receptor leptin. Leptin là một hormone được tiết ra từ các tế bào mỡ và có một số ảnh hưởng đến sinh lý, nhưng nó đặc biệt quan trọng trong việc điều khiển cảm giác thèm ăn và sự cân bằng năng lượng. Receptor leptin là một thành viên của nhóm rất nhiều các receptor cytokine, nhóm các receptor này không có hoạt động enzyme nội tại nhưng chúng có thể truyền tín hiệu đến các enzyme liên quan. Đối với receptor leptin, một trong những đường dẫn tín hiệu xảy ra thông qua tyrosin kinase của nhóm janus kinase(JAK), JAK2. Receptor leptin tồn tại dưới dạng dimer (2 tiểu phần), và sự gắn của leptin với phần bên ngoài màng tế bào của receptor làm thay đổi cấu trúc của nó, khởi động quá trình phosphoryl hóa và hoạt hóa các phân tử JAK2 liên quan trong nội bào. Sự kích hoạt các phân tử JAK2 sau đó gây phosphoryl hóa các tyrosin còn lại ở receptor leptin - hình thành phức hợp JAK2 làm trung gian tín hiệu nội bào. Những tín hiệu nội bào này bao gồm sự phosphoryl hóa của các protein chuyển đổi tín hiệu và kiểm soát phiên mã (STAT), kích hoạt quá trình phosphoryl hóa tại các gen leptin đích để bắt đầu quá trình tổng hợp protein. Sự phosphoryl hóa JAK2 còn dẫn đến hoạt hoá một số con đường enzyme nội bào khác như các kinase hoạt hóa phân bào (MAPK) hay phosphatidylinositol 3-kinase (PI3K). Một số ảnh hưởng của leptin diễn ra nhanh chóng là hậu quả của sự kích hoạt những enzyme nội bào này, ngược lại một số tác dụng khác diễn ra chậm hơn và cần phải có sự tổng hợp của những protein khác.
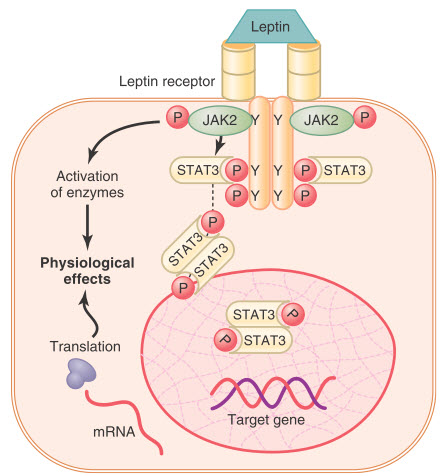
Hình. Thụ thể liên kết với enzym - thụ thể leptin. Thụ thể tồn tại dưới dạng homodimer (hai phần giống nhau), và leptin liên kết với phần ngoại bào của thụ thể, gây ra quá trình phosphoryl hóa (P) và kích hoạt janus kinase 2 liên kết nội bào (JAK2). Cơ chế này gây ra quá trình phosphoryl hóa chất dẫn truyền tín hiệu và chất hoạt hóa protein phiên mã (STAT), sau đó kích hoạt quá trình phiên mã của gen đích và tổng hợp protein. Quá trình phosphoryl hóa JAK2 cũng kích hoạt một số hệ thống enzym khác làm trung gian cho một số tác dụng nhanh hơn của leptin. Y, các vị trí phosphoryl hóa tyrosine cụ thể.
Một ví dụ khác, một cơ chế được sử dụng rộng rãi để điều khiển chức năng tế bào thông qua hormone, là cho hormone gắn với một receptor xuyên màng đặc biệt, receptor này sau đó sẽ trở thành enzyme hoạt hóa adenylyl cyclase nhô ra từ trong tế bào. Enzym này xúc tác sự hình thành cAMP, chất gây rất nhiều ảnh hưởng trong môi trường nội bào để kiểm soát các hoạt động của tế bào (sẽ được nói đến sau). cAMP được gọi là chất truyền tin thứ hai bởi vì bản thân đây không phải là một hormone trực tiếp gây ra sự thay đổi ở tế bào; thay vào đó, cAMP có chức năng như một chất truyền tin thứ hai để tạo ra những ảnh hưởng ở tế bào.
Với một vài hormone dạng peptid, như hormone lợi niệu tâm nhĩ (ANP), GMP vòng (cGMP), những chất chỉ khác cAMP một chút, cũng là những chất truyền tin thứ hai.
Những receptor hormone nội bào và sự hoạt hóa các gen
Có một vài hormone, bao gồm các hormone steroid tuyến thượng thận và tuyến sinh dục, hormone tuyến giáp, hormone vitamin A, vitamin D, gắn với các receptor protein bên trong tế bào nhiều hơn là gắn vào màng tế bào. Bời vì những hormone này tan trong dầu, chúng dễ dàng đi qua màng tế bào và gắn với các receptor trong tế bào chất hoặc nhân. Phức hợp hormone-receptor hoạt hóa sau đó gắn với một vùng đặc biệt (gọi là promoter) trên DNA và được gọi là giai đoạn đápứng hormone (hormone response element), và theo cách này thì hoặc sẽ kích hoạt hoặc ức chế phiên mã của các gen đặc hiệu và các ARN thông tin (mRNA, HÌNH 75-6). Do đó, vài phút, vài giờ hoặc thậm chí vài ngày sau khi hormone đi vào trong tế bào, một dạng protein mới xuất hiện trong tế bào và trở thành chất điều khiển các chức năng mới hoặc các chức năng khác của tế bào.
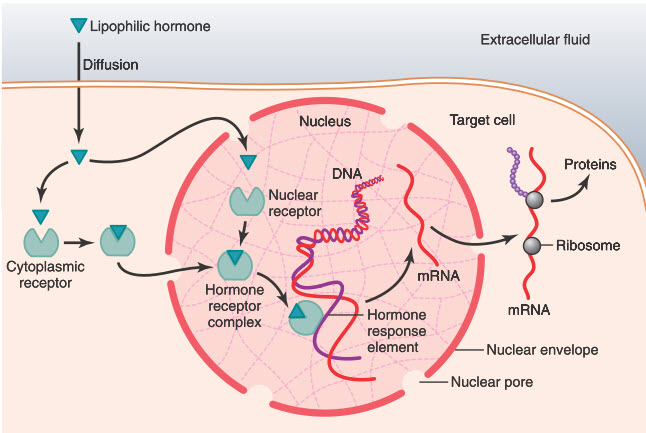
Hình. Cơ chế tương tác của hormone kỵ nước, chẳng hạn như steroid, với các thụ thể nội bào trong tế bào đích. Sau khi hormone liên kết với thụ thể trong tế bào chất hoặc trong nhân, phức hợp hormone-thụ thể sẽ liên kết với yếu tố đáp ứng hormone (promoter) trên DNA. Hành động này kích hoạt hoặc ức chế phiên mã gen, hình thành RNA thông tin (mRNA) và tổng hợp protein.
Nhiều loại mô khác nhau có cùng loại receptor nội bào giống nhau, tuy nhiên gen được kích thích bởi các receptor ở một số mô lại khác nhau. Một receptor nội bào có thể kích hoạt một gen chỉ khi có sự liên kết phù hợp giữa receptor với protein điều hòa gen, và nhiều loại pritein này là đặc hiệu cho từng loại mô cơ thể. Do đó, đáp ứng của các loại mô khác nhau với hormone được tìm hiểu không chỉ về tính chất đặc hiệu của các receptor mà còn cả biểu hiện của gen được receptor điều khiển.
Bài viết cùng chuyên mục
Chức năng tạo nước tiểu sinh lý của thận
Cầu thận được cấu tạo bởi một mạng lưới mao mạch, xếp song song, và được bao quanh bởi bao Bowman.
Dinh dưỡng cho phôi
Khi các tế bào lá nuôi phôi tràn vào màng rụng, tiêu hoá và hấp thu nó, các chất dinh dưỡng được lưu trữ trong màng rụng được phôi sử dụng cho sự sinh trưởng và phát triển.
Lưu lượng máu đến ruột chịu ảnh hưởng của hoạt động và tác nhân chuyển hóa ruột
Mặc dù các nguyên nhân chính xác gây tăng lượng máu khi đường tiêu hóa hoạt động vẫn chữa được biết rõ, nhưng một vài phần đã được làm sáng tỏ.
Vai trò và chức năng của Protein huyết tương
Proteins huyết tương là một nguồn amio acid của mô, khi các mô cạn kiệt protein, các protein huyết tương có thể hoạt động như một nguồn thay thế nhanh chóng.
Đám rối thần kinh cơ ruột và đám rối thần kinh dưới niêm mạc
Đám rối thần kinh cơ ruột không hoàn toàn có tác dụng kích thích vì một số neuron của nó có tác dụng ức chế; tận cùng của các sợi đó tiết ra một chất ức chế dẫn truyền, có thể là “polypeptide hoạt mạch ruột”.
Cấu trúc chức năng sinh lý tim
Thành cơ tim thất trái dày gấp hai đến bốn lần thành thất phải, do nó phải bơm máu với áp lực cao hơn để thắng sức cản lớn của tuần hoàn hệ thống.
Các sợi thần kinh cảm giác hướng tâm từ ruột
Có nhiều sợi thần kinh cảm giác hướng tâm phân bố tại ruột. Một số sợi thần kinh có thân neuron nằm trong hệ thần kinh ruột và một số lại nằm trong hạch gai của tủy sống.
Hệ tuần hoàn: dòng máu chảy
Tất cả lưu lượng máu trong toàn bộ hệ tuần hoàn ở người trưởng thành khoảng 5000ml/phút. Đó là lượng lưu lượng tim bởi vì lượng máu bơm qua động mạch bởi tim trong mỗi phút.
Hệ mạch cửa dưới đồi yên của thùy trước tuyến yên
Mạch máu đi vào các xoang đầu tiên đều đi qua giường mao mạch ở phần dưới vùng dưới đồi, dòng máu sau đó chảy qua các mạch cửa dưới đồi yên rồi đổ vào các xoang ở tuyến yên trước.
Dược lý của hệ thần kinh tự chủ
Các thuốc có tác dụng gián tiếp lên hệ giao cảm tại vị trí các cơ quan đích của hệ giao cảm bị kích thích trực tiếp. Bao gồm các thuốc ephedrine, tyramine, và amphetamine.
Tổng hợp các Triglycerides từ Carbohydrates
Tổng hợp triglycerides xảy ra ở gan, nhưng một lượng nhỏ cũng được tổng hợp ở mô mỡ. Triglycerides được tổng hợp ở gan sẽ được vận chuyển chủ yếu dưới dạng VLDLs tới mô mỡ, nơi chúng được dự trữ.
Sự tăng trưởng và phát triển chức năng của bào thai
Do trọng lượng thai tương ứng xấp xỉ với lập phương của chiều dài, trọng lượng thai hầu như tăng tương ứng với lập phương tuổi thai.
Các sợi thần kinh: dẫn truyền các loại tín hiệu khác nhau và phân loại
Các sợi thần kinh đến có kích thước trung bình vào khoảng 0.5 đến 20 micromet; kích thước lớn hơn, tốc độ dẫn truyền cũng lớn hơn. Giới hạn tốc độ dẫn truyền trong khoảng 0.5 đến 120 m/giây.
Sinh lý quá trình sinh nhiệt thải nhiệt cơ thể
Hầu hết lượng nhiệt sinh ra trong cơ thể được tạo thành từ các cơ quan ở sâu như gan, não, tim và cơ (khi có vận cơ). Rồi thì nhiệt được vận chuyển đến da là nơi có thể thải nhiệt vào môi trường xung quanh.
Sinh lý tuần hoàn địa phương (mạch vành, não, phổi)
Động mạch vành xuất phát từ động mạch chủ, ngay trên van bán nguyệt. Động mạch vành đến tim, chia thành động mạch vành phải và trái. Động mạch vành trái cung cấp máu cho vùng trước thất trái và nhĩ trái.
Chuyển hóa khoáng chất trong cơ thể
Số lượng trong cơ thể của các khoáng chất quan trọng nhất, và nhu cầu hằng ngày được cung cấp gồm magnesium, calcium, phosphorus, sắt, những nguyên tố vi lượng.
Ảnh hưởng của gia tốc tuyến tính lên cơ thể
Vấn đề cũng xẩy ra trong quá trình giảm tốc, khi tàu không gian trở vào khí quyển. Một người di chuyển ở Mach 1 (tốc độ âm thanh và của máy bay nhanh) có thể giảm tốc an toàn trong khoảng cách tầm 0.12 dặm.
Chức năng của hệ limbic: vị trí chủ chốt của vùng dưới đồi
Cấu trúc giải phẫu của hệ limbic, cho thấy phức hợp kết nối của các thành phần nội liên kết trong não. Nằm ở giữa những cấu trúc này là vùng dưới đồi, kích thước vô cùng nhỏ.
Điện thế hoạt động của tế bào thần kinh
Để tạo ra những tín hiệu thần kinh, điện thế hoạt động di chuyển dọc theo tế bào sợi thần kinh cho tới điểm kếtthúc của nó.
Điểm nhiệt chuẩn trong điều nhiệt cơ thể
Điểm nhiệt chuẩn tới hạn của vùng dưới đồi, mà ở đó ở dưới mức run cơ và trên mức bắt đầu đổ mồ hôi, được xác định chủ yếu bởi hoạt động của receptor càm nhận nhiệt trong vùng trước thị-trước dưới đồi.
Vận chuyển lipids trong dịch cơ thể
Cholesterol và phospholipid được hấp thụ từ hệ thống ruột vào trong chylomicron. Vì thế dù chylomicron được cấu tạo chủ yếu từ triglycerides, chúng còn chứa phospholipid, cholesterol và apoprotein B.
Tác dụng lên thận và tuần hoàn của Aldosterol
Aldosterol làm tăng tái hấp thu natri và đồng thời tăng bài tiết kali trong các tế bào chính của ống thận nhỏ nhưng cũng bài tiết ion hydro để trao đổi với kali vào trong tế bào vỏ ống góp.
Sinh lý thần kinh hành não
Hành não là phần thần kinh trung ương tiếp nối với tủy sống, nằm ở phần thấp nhất của hộp sọ, ngay sát trên lỗ chẩm. Hành não là nơi xuất phát của nhiều dây thần kinh sọ (từ dây V đến dây XII) trong đó quan trọng nhất là dây X.
Sinh lý hệ mạch máu
Tim trái tống máu vào động mạch chủ, tạo ra một áp lực lớn đưa máu qua vòng tuần hoàn cho đến tim phải: áp lực cao nhất trong động mạch chủ và thấp nhất trong tâm nhĩ phải.
Phát triển của phôi trong tử cung
Khi sự cấy diễn ra, các tế bào lá nuôi phôi và các tế bào lá nuôi lân cận (từ túi phôi và từ nội mạc tử cung) sinh sản nhanh chóng, hình thành nhau thai và các màng khác nhau của thai kì, phôi nang.
