- Trang chủ
- Sách y học
- Sinh lý y học
- Đặc điểm phân tử của các sợi cơ co bóp
Đặc điểm phân tử của các sợi cơ co bóp
Một đặc tính của đầu myosin mà cần thiết cho sự co cơ là nó có chức năng như một enzyme adenosine triphosphatase (ATPase).
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Các sợi myosin là bao gồm của các phân tử myosin kép
Mỗi một phân tử myosin, thể hiện trong hình A, có trọng lượng phân tử khoảng 480.000. Hình B cho thấy tổ chức của nhiều phân tử để tạo thành một sợi myosin, cũng như sự tương tác của sợi này trên một mặt với các đầu tận của hai sợi actin.
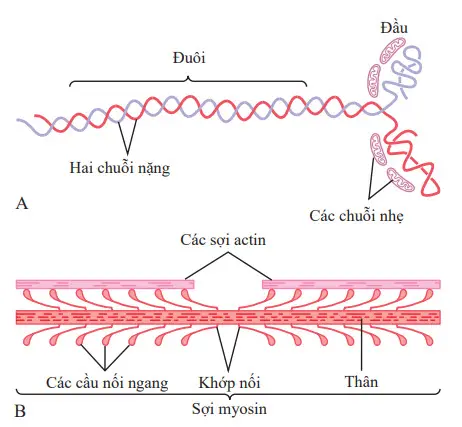
Hình. A, Phân tử myosin. B, Sự kết hợp của nhiều phân tử myosin tạo thành một sợi myosin. Cũng được thể hiện là hàng ngàn các cầu nối chéo myosin và sự tương tác giữa các đầu của các cầu nối chéo với các sợi actin liền kề.
Phân tử myosin (xem hình A) bao gồm sáu chuỗi polypeptide - hai chuỗi nặng, mỗi chuỗi có trọng lượng phân tử khoảng 200.000, và bốn chuỗi nhẹ với trọng lượng phân tử khoảng 20.000 mỗi chuỗi. Hai chuỗi nặng quấn xoắn quanh nhau để tạo thành một chuỗi xoắn kép, được gọi là đuôi của phân tử myosin. Một đầu của mỗi chuỗi được gấp song phương thành một cấu trúc polypeptide hình cầu gọi là một đầu myosin.
Như vậy, có hai đầu tự do ở một đầu của chuỗi xoắn kép phân tử myosin. Bốn chuỗi nhẹ cũng là một phần của đầu myosin, hai ở mỗi đầu. Các chuỗi nhẹ này giúp kiểm soát chức năng của đầu trong quá trình co cơ.
Sợi myosin được tạo thành từ 200 hoặc nhiều hơn các phân tử myosin riêng lẻ. Phần trung tâm của một trong các sợi này được thể hiện trong hình hình B, sự xuất hiện các đuôi của các phân tử myosin bó lại với nhau để hình thành phần thân của sợi, trong khi nhiều đầu của các phân tử treo bên ngoài đến các bên của thân. Ngoài ra, một phần của thân của mỗi phân tử myosin treo đến bên cạnh cùng với đầu, do đó cấp một nhánh gửi tới đầu ngoài từ thân, như thể hiện trong hình. Các nhánh nhô ra và các đầu cùng nhau được gọi là các cầu nối chéo. Mỗi cầu nối chéo linh hoạt ở hai điểm gọi là khớp nối-một trong những nơi nhánh rời khỏi thân của sợi myosin, và khác nơi mà đầu gắn vào nhánh. Các nhánh có khớp nối cho phép các đầu được hoặc kéo dài xa ra ngoài từ thân của sợi myosin hoặc đưa lại gần tới thân. Các đầu co khớp nối lần lượt tham gia vào quá trình co bóp thực tế, như thảo luận trong các phần sau đây.
Tổng chiều dài của mỗi sợi myosin là không đổi - gần như chính xác 1,6µm. Lưu ý, tuy nhiên, không có cầu nối ngang các đầu ở trung tâm của sợi myosin cho một khoảng cách khoảng 0,2µm vì các nhánh có khớp nối kéo dài đi xa từ trung tâm.
Bây giờ, để hoàn thành bức tranh, sợi myosin xoắn lại do đó mỗi cặp liên tiếp của các cầu nối chéo có trục di dời từ cặp trước đó bằng 120 độ. Việc xoắn này đảm bảo rằng các cầu nối ngang kéo dài ở tất cả các hướng xung quanh sợi.
Adenosine Triphosphatase hoạt động của đầu myosin
Một đặc tính khác của đầu myosin mà cần thiết cho sự co cơ là nó có chức năng như một enzyme adenosine triphosphatase (ATPase). Như giải thích sau, đặc tính này cho phép đầu tách ra ATP và sử dụng năng lượng có nguồn gốc từ liên kết cao năng phosphat của ATP để nạp năng lượng cho quá trình co bóp.
Các sợi Actin bao gồm có actin, tropomyosin, và troponin
Trụ cột của sợi actin là một phân tử protein F-actin sợi kép, được miêu tả bởi hai sợi màu sáng hơn trong hình. Hai sợi được quấn trong một chuỗi xoắn theo cách tương tự như phân tử myosin.
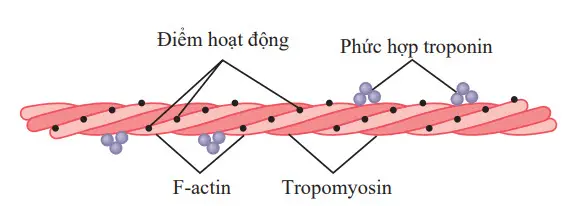
Hình. Sợi Actin, bao gồm hai chuỗi xoắn ốc của các phân tử F-actin và hai sợi phân tử tropomyosin phù hợp với các rãnh giữa các sợi Actin. Được gắn vào một đầu của mỗi phân tử tropomyosin là một phức hợp troponin bắt đầu sự co lại.
Mỗi sợi của chuỗi xoắn kép F-actin bao gồm các phân tử G-actin polyme hóa, mỗi cái có một trọng lượng phân tử khoảng 42.000. Kèm theo mỗi một của các phân tử G-actin là một phân tử ADP. Các phân tử ADP này được tin là các điểm hoạt động trên các sợi actin mà cùng các cầu nối chéo của các sợi myosin tương tác để gây co cơ. Các điểm hoạt động trên hai sợi F-actin của chuỗi xoắn kép xen kẽ nhau, cho một điểm hoạt động trên toàn bộ sợi actin là khoảng 2,7nm mỗi điểm.
Mỗi sợi actin dài khoảng 1µm. Nền của các sợi actin được chèn mạnh vào trong các đĩa Z; các đầu của các sợi nhô ra ở cả hai chiều nằm trong không gian giữa các các phân tử myosin, như thể hiện trong hình.
Các phân tử Tropomyosin
Sợi actin cũng chứa một loại protein khác, tropomyosin. Mỗi phân tử của tropomyosin có trọng lượng phân tử là 70.000 và chiều dài là 40 nm.
Những phân tử này được quấn xoắn xung quanh các mặt của chuỗi xoắn F-actin. Ở trạng thái nghỉ, các phân tử tropomyosin nằm trên đầu của các điểm hoạt động của các sợi actin bởi vậy không thể xảy ra thu hút giữa các sợi actin và myosin để gây ra co bóp.
Troponin và vai trò của nó trong sự co cơ
Gắn không liên tục dọc theo hai bên của các phân tử tropomyosin là những phân tử protein bổ sung được gọi là troponin. Các phân tử protein này thực chất là phức hợp của ba tiểu đơn vị protein liên kết lỏng lẻo, mỗi một trong số đó đóng một vai trò đặc thù trong việc kiểm soát sự co cơ.
Một trong những tiểu đơn vị (troponin I) có ái lực mạnh với actin, tiểu đơn vị khác (troponin T) với tropomyosin, và cái thứ ba (troponin C) với các ion canxi. Phức hợp này được cho là gắn tropomyosin với actin. Ái lực mạnh của troponin với các ion canxi được cho là khởi đầu quá trình co bóp, như được giải thích trong phần tiếp theo.
Sự tương tác của một sợi myosin, hai sợi actin, và các ion canxi để gây ra co bóp.
Sự ức chế của sợi Actin bởi phức hợp Troponin – Tropomyosin
Một actin sợi hoàn toàn không có sự hiện diện của phức hợp troponin-tropomyosin (nhưng có sự hiện diện của các ion magiê và ATP) liên kết ngay lập tức và mạnh với các đầu của các phân tử myosin. Sau đó, nếu phức hợp troponin-tropomyosin được thêm vào các sợi actin, sự liên kết giữa myosin và actin không diễn ra. Do đó, người ta tin rằng các điểm hoạt động trên sợi actin bình thường của cơ giãn là bị ức chế hoặc bị che phủ vị trí bởi phức hợp troponintropomyosin. Do đó, các điểm không thể gắn với các đầu của các sợi myosin để gây ra co bóp. Trước khi co bóp có thể xảy ra, tác dụng ức chế của chính phức hợp troponin-tropomyosin phải bị ức chế.
Sự hoạt hóa của sợi Actin bởi các ion canxi
Trong sự hiện diện của một lượng lớn của các ion canxi, tác dụng ức chế của troponin-tropomyosin trên các sợi actin chính nó lại bị ức chế. Cơ chế của sự ức chế này chưa được biết, nhưng có một giả thiết như sau: Khi các ion canxi kết hợp với troponin C, mỗi phân tử có thể liên kết mạnh với tối đa bốn ion canxi, phức hợp troponin được cho là trải qua một sự thay đổi về hình dạng mà trong một số cách kéo trên phân tử tropomyosin và di chuyển sâu hơn vào các rãnh giữa hai sợi actin. Hoạt động này “bộc lộ” các vị trí hoạt động của actin, do đó cho phép các vị trí hoạt động này thu hút các đầu của cầu nối chéo myosin và khiến sự co bóp được tiến hành. Mặc dù cơ chế này là giả thuyết, nó nhấn mạnh rằng mối quan hệ bình thường giữa phức hợp troponintropomyosin và actin bị thay đổi bởi các ion canxi, sinh ra một điều kiện mới dẫn đến sự co bóp.
Sự tương tác của sợi Actin “hoạt hóa” và các cầu nối chéo Myosin - Lý thuyết “đi bộ dọc” của sự co bóp
Ngay sau khi sợi actin được hoạt hóa bởi các ion canxi, đầu của các cầu nối chéo từ các sợi myosin trở nên thu hút với các vị trí hoạt động của sợi actin, và điều này, theo cách nào đó, khiến sự co bóp xảy ra. Mặc dù cách thức chính xác mà sự tương tác giữa các cầu nối chéo và actin gây ra sự co bóp vẫn còn phần nào là lý thuyết, một giả thuyết mà có bằng chứng đáng kể tồn tại là lý thuyết “đi bộ dọc” (hoặc “chốt cài”) của sự co bóp.
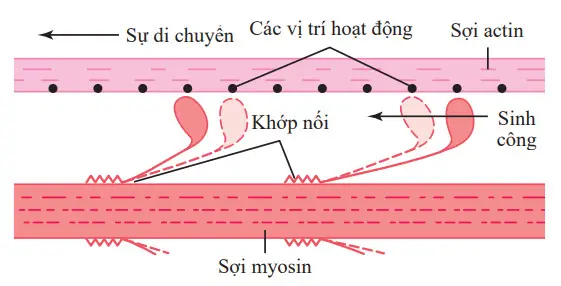
Hình. Cơ chế “đi bộ dọc” cho sự co bóp của cơ
Hình chứng tỏ giả thiết cơ chế đi bộ dọc với sự co bóp. Hình cho thấy đầu của hai cầu nối chéo gắn vào và tách ra khỏi các vị trí hoạt động của một sợi actin.
Khi một đầu gắn vào một vị trí hoạt động, sự liên kết này đồng thời gây ra những thay đổi sâu sắc trong năng lượng nội phân tử giữa đầu và nhánh cầu chéo của nó.
Sự điều chỉnh mới của năng lượng làm cho đầu nghiêng về phía nhánh và kéo theo sợi actin cùng với nó. Sự nghiêng này của đầu được gọi là sinh công (power stroke). Ngay sau khi nghiêng, đầu sau đó tự động tách ra khỏi vị trí hoạt động. Tiếp theo, đầu trở lại hướng kéo dài của nó. Ở vị trí này, nó kết hợp với một vị trí hoạt động mới xa hơn xuống dọc theo sợi actin; đầu sau đó nghiêng một lần nữa để gây ra một sinh công mới, và sợi actin di chuyển thêm một bước. Do đó, đầu của các cầu chéo uốn cong trở lại và về phía trước và từng bước đi bộ dọc sợi actin, kéo hai đầu của hai sợi actin liên tiếp về phía trung tâm của sợi myosin.
Mỗi một của các cầu nối chéo được cho là hoạt động độc lập với tất cả các cầu nối khác, từng sự gắn và kéo trong một chu kỳ lặp đi lặp lại liên tục. Vì vậy, số cầu nối chéo tiếp xúc với sợi actin tại bất kỳ thời điểm nhất định nào càng lớn, lực của co bóp càng lớn.
ATP như nguồn năng lượng cho sự co bóp - Các sự kiện hóa học trong chuyển động của các đầu Myosin
Khi một cơ co, làm việc được thực hiện và năng lượng là cần thiết. Một lượng lớn ATP được tách ra để tạo thành ADP trong suốt quá trình co bóp, và khối lượng công việc được thực hiện bởi cơ càng nhiều, lượng ATP được tách ra càng nhiều; hiện tượng này được gọi là hiệu ứng Fenn. Các sự kiện nối tiếp dưới đây được cho là phương pháp để mà hiệu ứng này xảy ra:
1. Trước khi sự co bóp bắt đầu, đầu của các cầu chéo gắn với ATP. ATPase hoạt động của đầu myosin ngay lập tức sẽ tách ATP nhưng để lại sản phẩm tách, ADP cộng ion phosphate, liên kết với đầu.
Trong trạng thái này, hình dáng của đầu bởi vậy mà nó mở rộng đường vuông góc về phía sợi actin nhưng vẫn chưa gắn vào actin.
2. Khi phức hợp troponin-tropomyosin liên kết với các ion canxi, các vị trí hoạt động trên sợi actin được bộc lộ và các đầu myosin sau đó liên kết với các vị trí này, như thể hiện trong hình.
3. Sự liên kết giữa đầu của các cầu nối chéo và vị trí hoạt động của sợi actin gây ra một sự thay đổi về hình dạng tại đầu, khiến đầu nghiêng về phía nhánh của cầu nối chéo và thực hiện sinh công cho sự kéo sợi actin. Năng lượng mà kích hoạt sinh công là năng lượng đã được dự trữ, giống như một lò xo “vểnh lên”, bởi sự thay đổi về hình dạng mà đã xảy ra trong đầu khi phân tử ATP được tách ra trước đó.
4. Một khi đầu của cầu nối chéo nghiêng, sự giải phóng của ADP và ion phosphate mà trước đó gắn vào đầu được cho phép. Tại vị trí của giải phóng của ADP, một phân tử mới của ATP liên kết. Liên kết này của ATP mới gây ra tách rời của đầu khỏi actin.
5. Sau khi đầu đã tách ra khỏi actin, phân tử mới của ATP được tách ra để bắt đầu cho chu kỳ tiếp theo, dẫn tới một sinh công mới. Đó là, năng lượng “vểnh lên” lần nữa để đầu trở về trạng thái vuông góc của nó, sẵn sàng để bắt đầu chu kỳ sinh công mới.
6. Khi đầu vểnh lên (với năng lượng dự trữ của nó bắt nguồn từ ATP tách ra) liên kết với một vị trí hoạt động mới trên sợi actin, nó trở nên không vểnh lên và một lần nữa thực hiện một sinh công mới.
Như vậy, quá trình được tiến hành hết lần này đến lần khác cho đến khi các sợi actin kéo màng Z gần sát với các đầu của các sợi myosin hoặc cho đến khi mức tải trên cơ trở nên quá lớn để có thêm sự kéo xảy ra.
Bài viết cùng chuyên mục
Đặc điểm sinh lý cấu tạo bộ máy tiêu hóa
Thiết đồ cắt ngang, ống tiêu hóa cấu tạo gồm 8 lớp từ trong ra ngoài: niêm mạc, cơ niêm, lớp dưới niêm mạc, đám rối Meissner, cơ vòng
Giải phẫu sinh lý thành ống tiêu hóa
Thành ruột, từ ngoài vào trong bao gồm các lớp sau đây: lớp thanh mạc, lớp cơ trơn dọc, lớp cơ trơn vòng, lớp dưới niêm mạc, và lớp niêm mạc. Thêm vào đó, có rải rác các sợi cơ trơn nằm sâu ở lớp niêm mạc được gọi là lớp cơ niêm.
Sự tiết Progesterone của nhau thai
Progesterone góp phần vào sự phát triển của thai, làm tăng bài tiết của ống dẫn trứng và tử cung người mẹ ddeer cung cấp chất dinh dưỡng thích hợp cho sự phát triển của phôi dâu và túi phôi.
Chức năng của Lipoproteins trong vận chuyển Cholesterol và Phospholipids
Hầu hết các lipoprotein được hình thành ở gan, đó cũng là nơi mà hầu hết các cholesterol huyết tương, phospholipid và triglycerides được tổng hợp.
Block nút nhĩ thất: chặn đường truyền tín hiệu điện tim
Thiếu máu nút nhĩ thất hoặc bó His thường gây chậm hoặc block hẳn dẫn truyền từ nhĩ đến thất. Thiếu máu mạch vành có thể gây ra thiếu máu cho nút nhĩ thất và bó His giống với cơ chế gây thiếu máu cơ tim.
Sự xâm nhập tinh trùng vào trứng: các enzyme thể đỉnh và phản ứng thể đỉnh
Chỉ vài phút sau khi tinh trùng đầu tiên xâm nhập qua lớp màng sáng, các ion calci xâm nhập vào bên trong, tế bào phát động phản ứn g vỏ đổ các chất đặc biệt vào xoang quanh noãn.
Dẫn truyền thị giác: đường dẫn truyền từ hai võng mạc đến vỏ não thị giác
Đường dẫn thị giác có thể được chia sơ bộ thành một hệ thống cũ tới trung não và nền não trước và một hệ thống mới để truyền trực tiếp tín hiệu hình ảnh về vỏ não thị giác ở thùy chẩm.
Điều chỉnh lượng thức ăn ăn vào và dự trữ năng lượng của cơ thể
Duy trì sự cung cấp năng lượng đầy đủ trong cơ thể quan trọng đến nỗi mà rất nhiều các cơ chế kiểm soát ngắn hạn và dài hạn tồn tại không chỉ điều chỉnh năng lượng hấp thu mà cả năng lượng tiêu thụ và năng lượng dự trữ.
Hô hấp trong tập luyện thể thao
Có một mối quan hệ tuyến tính. Cả tiêu thụ oxy và tổng thông khí phổi tăng gấp khoảng 20 lần từ trạng thái nghỉ ngơi và cường độ tập luyện tối đa ở các vận động viên được tập luyện tốt.
Kích thích và dẫn truyền xung động của tim
Nút xoang (còn gọi là nhĩ xoang hay nút SA) phát nhịp trong hệ thống tạo xung nhịp bình thường, theo đường dẫn xung từ nút xoang tới nút nhĩ thất (AV).
Sự vận động của tế bào cơ thể người
Yếu tố cần thiết của sự chuyển động là cung cấp năng lượng cần thiết để kéo tế bào về phía chân giả. Trong bào tương của tất cả tế bào là một lượng lớn protein actin.
Khoảng các từ vật tới mắt: xác định chiều sâu
Nếu ta đã biết một người nào đó cao 6 feet, ta có thể biết được người đó cách ta bao xa thông qua kích thước ảnh của người đó trên võng mạc.
Điều hòa lưu lượng máu bằng cách phát triển tuần hoàn bàng hệ
Sự mở các tuần hoàn bàng hệ sau đó trong vòng nhiều giờ kế tiếp, sao cho trong vòng 1 ngày, một nửa mô cần máu có thể được đáp ứng, và trong vòng 1 vài ngày dòng máu thường đủ để đến các mô.
Điều hòa gen trong cơ thể người
Ở động vật có nhiều loại tế bào, mô, cơ quan khác nhau, các điều hòa biểu hiện gen khác nhau cũng cho phép nhiều loại tế bào khác nhau trong cơ thể thực hiện các chức năng chuyên biệt của chúng.
Hormone parathyroid (tuyến cận giáp)
Tuyến cận giáp của con người trưởng thành, chứa chủ yếu là các tế bào chính và một số lượng nhỏ đến trung bình các tế bào oxyphil, nhưng tế bào oxyphil vắng mặt ở nhiều loài động vật cũng như ở người trẻ.
Chức năng của Mineralocorticoids Aldosterone
Aldosterol chiếm phần lớn hoạt tính mineralocorticoid của hormon vỏ thượng thận, nhưng corticoid là glucocorticoid chính được tiết ở vỏ thượng thận, cũng tham giá đáng kể vào hoạt tính của mineralocorticoid.
Điện thế hoạt động trong cơ tim
Trong cơ tim, điện thế hoạt động được tạo ra do mở kênh natri nhanh kích hoạt điện thế và một tập hợp hoàn toàn khác các kênh canxi typ L, chúng được gọi là kênh canxi - natri.
Ngoại tâm thu nhĩ: rối loạn nhịp tim
Khi tim co sớm hơn bình thường, tâm thất chưa nhận đầy máu như bình thường và nhát bóp đó bơm ít máu hơn. Do đó sóng đập của nhát bóp đó lên thành mach sẽ yếu hơn thậm chí là yếu đến mức không thể bắt được gọi làm mạnh chìm.
Chức năng sinh dục nam bất thường
Rối loạn chức năng cương dương, hay gọi là “bất lực”, đặc trưng bởi sự mất khả năng duy trì độ cương cứng của dương vật để thực hiện quá trình giao hợp phù hợp.
Sự tiết estrogen của nhau thai
Estrogen chủ yếu gây ra một sự tăng sinh trên hầu hết các cơ quan sinh sản và liên quan đến người mẹ. Trong khi mang thai estrogen làm cho tử cung được mở rộng, phát triển vú và ống vú người mẹ, mở rộng cơ quan sinh dục ngoài.
Tái tạo mạch máu để đáp ứng với những thay đổi mãn tính về lưu lượng hoặc áp lực máu
Khi áp lực dòng máu cao trường kì hơn mức bình thường, các động mạch và tiểu động mạch lớn nhỏ cấu trúc lại để thành mạch máu thích nghi với áp lực mạch máu lớn hơn.
Nút xoang (xoang nhĩ): hệ thống kích thích và dẫn truyền của tim
Nút xoang nhỏ, dẹt, hình dải elip chuyên biệt của cơ tim rộng khoảng 3mm, dài 15mm và dày 1mm. Nó nằm ở sau trên vách tâm nhĩ phải, ngay bên dưới và hơi gần bên chỗ mở của tĩnh mạch chủ trên.
Rung thất: cơ chế phản ứng dây truyền rối loạn nhịp tim
Vòng đầu tiên của kích điện tim gây ra sóng khử cực lan mọi hướng, khiến cho tất cả cơ tim đều ở trạng thái trơ. Sau 25s. một phần của khối cơ này thoát khỏi tình trạng trơ.
Hoạt động chuyển hóa tế bào của Aldosterol
Aldosterol không có tác dụng vận chuyển natri ngay lập tức; đúng hơn tác dụng này phải chờ hình thành liên tục những chất đặc biệt cần thiết vận chuyển natri.
Điều hòa sự bài tiết hormone tăng trưởng (GH)
Cơ chế chính xác điều khiển sự bài tiết GH vẫn chưa được hiểu một cách hoàn toàn, nhưng có vài yếu tố liên quan tới mức độ dinh dưỡng của cơ thể hoặc căng thẳng đã được biết là các yếu tố gây kích thích bài tiết GH.
