- Trang chủ
- Sách y học
- Sinh lý y học
- Sự giải phóng năng lượng từ Glucose cho cơ thể theo con đường đường phân
Sự giải phóng năng lượng từ Glucose cho cơ thể theo con đường đường phân
Cách quan trọng nhất để giải phóng năng lượng từ glucose là khởi động con đường đường phân, sản phẩm cuối cùng sau đó được oxy hóa để cung cấp năng lượng.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Vì sự oxy hóa hoàn toàn 1 mol glucose giải phóng ra 686,000 calo và chỉ cần 12,000 calo để tổng hợp nên 1 mol ATP, năng lượng có thể bị lãng phí nếu glucose bị phân hủy toàn bộ thành nước và CO2 trong khi chỉ tạo ra một phân tử ATP. May mắn là, tế bào trong cơ thể chứa những enzym đặc biệt làm cho phân tử glucose chia cắt từng phần nhỏ trong một thời điểm và qua nhiều bước liên tiếp, vì thế năng lượng được giải phóng ra thành từng phần nhỏ để tạo nên một phân tử ATP tại một thời điểm, do đó mỗi mol glucose được tế bào chuyển hóa tạo ra tổng cộng 38 mol ATP.
Đường phân (Glycolysis) - Glucose thành Pyruvic Acid
Cho đến nay cách quan trọng nhất để giải phóng năng lượng từ glucose là khởi động con đường đường phân. Sản phẩm cuối cùng của quá trình đường phân sau đó được oxy hóa để cung cấp năng lượng. Đường phân nghĩa là chia cắt một phân tử glucose để tạo thành hai phân tử pyruvic acid.
Đường phân xảy ra bởi 10 phản ứng hóa học liên tiếp. Mỗi bước được xúc tác bởi ít nhất một enzym đặc hiệu. Chú ý rằng ban đầu, glucose được chuyển thành fructose-1,6-diphosphate và sau đó chia thành hai phân tử có 3-carbon , glyceraldehyde-3-phosphate, chúng sẽ được chuyển thành pyruvic acid thông qua 5 bước nữa.
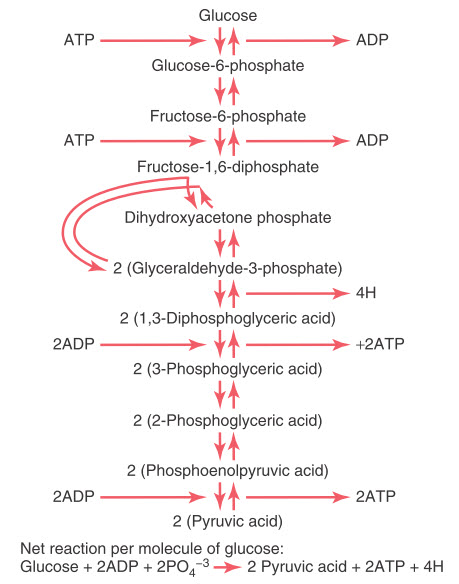
Hình. Chuỗi các phản ứng hóa học gây ra quá trình đường phân.
Sự tạo thành ATP trong quá trình đường phân. Mặc dù có nhiều phản ứng hóa học trong quá trình đường phân, chỉ có một phần nhỏ năng lượng tự do trong phân tử glucose được giải phóng tại hầu hết các bước. Mặc dù vậy, trong giai đoạn giữa 1,3-diphosphoglyceric acid và 3-phosphoglyceric acid, và giai đoạn giữa phosphoenolpyruvic acid và pyruvic acid, một phần của năng lượng được giải phóng lớn hơn 12,000 calo mỗi mol, số lượng cần thiết để tổng hợp nên ATP, và phản ứng tổng hợp ATP được hình thành. Do đó, tổng cộng 4 mol ATP đã được tạo ra từ 1 mol fructose-1,6-diphosphate được phân chia thành pyruvic acid.
Tuy nhiên, cần 2 mol ATP để phosphoryl hóa glucose ban đầu thành fructose-1,6-diphosphate trước khi quá trình đường phân bắt đầu. Do đó, số lượng thực tế thu được của cả quá trình đường phân chỉ là 2 mol ATP cho mỗi mol glucose được sử dụng. Số lượng này tương đương 24000 calo đã được chuyển thành ATP, nhưng trong quá trình đường phân, tổng cộng có 56,000 calo đã mất từ phân tử glucose ban đầu, thành ra hiệu suất của toàn bộ quá trình tổng hợp ATP chỉ là 43%. 57% năng lượng còn lại mất đi dưới dạng nhiệt.
Chuyển hóa Pyruvic Acid thành Acetyl Coenzyme A
Giai đoạn tiếp theo trong sự thoái hóa của glucose là chuyển hóa qua hai bước của hai phân tử pyruvic acid thành hai phân tử acetyl coenzyme A (acetyl-CoA) bằng các phản ứng sau:
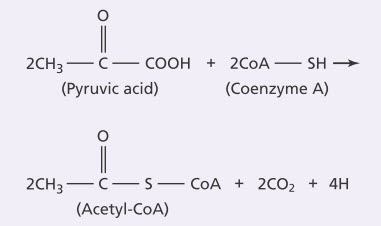
Hai phân tử CO2 và 4 nguyên tử hydro được giải phóng từ phản ứng này, trong khi phần còn lại của hai phân tử pyruvic acid liên kết với coenzyme A, là một dẫn xuất của vitamin B5 (pantothenic acid), để tạo thành hai phân tử acetyl-CoA. Trong chuyển hóa này, không có ATP nào được tạo ra, nhưng tới 6 phân tử ATP sẽ được tạo thành khi 4 nguyên tử hydro vừa giải phóng được oxy hóa sau đó và sẽ được đề cập sau đây.
Chu trình Citric Acid (Chu trình Krebs)
Giai đoạn tiếp theo của quá trình thoái hóa glucose gọi là chu trình citric acid (còn được biết dưới cái tên chu trình tricarboxylic acid hoặc chu trình Krebs nhằm vinh danh Hans Krebs - người đã khám phá ra chu trình này). Chu trình citric acid là một chuỗi các phản ứng hóa học trong đó phần acetyl của phân tử acetyl-CoA được thoái hóa thành CO2 và nguyên tử Hydro. Tất cả những phản ứng này đều xảy ra ở trong chất nền của ty thể. Sự giải phóng nguyên tử hydro làm tăng thêm số lượng nguyên tử sẽ bị oxy hóa, giải phóng ra một năng lượng cực lớn để hình thành nên ATP.
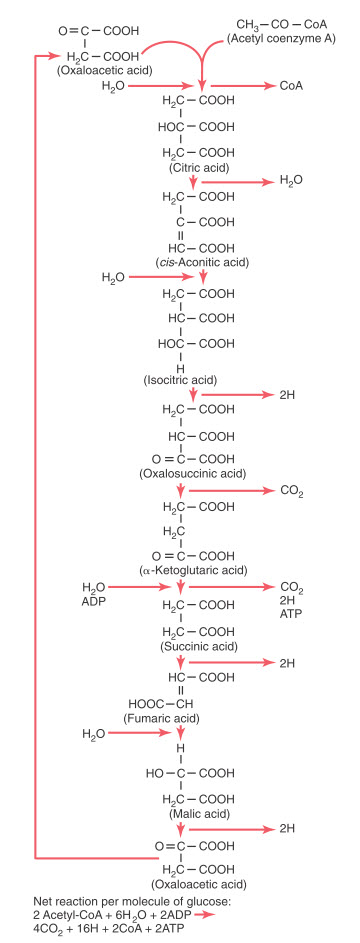
Hình. Phản ứng hóa học của chu trình axit xitric, cho thấy sự giải phóng khí cacbonic và một số nguyên tử hydro trong chu trình.
Các giai đoạn khác nhau của các phản ứng hóa học trong chu trình citric acid. Ở bên trái là các chất tham gia thêm vào phản ứng, còn sản phẩm của phản ứng được thể hiện ở bên phải. Chú ý rằng vị trí cao nhất trong sơ đồ là nơi chu trình bắt đầu với oxaloacetic acid, và khi chuỗi phản ứng kết thúc, oxaloacetic acid được tái tổng hợp một lần nữa. Do đó chu trình có thể được lặp lại nhiều lần.
Trong giai đoạn khởi đầu của chu trình citric acid, acetyl-CoA gắn với oxaloacetic acid tạo thành citric acid. Phần coenzyme A của acetyl-CoA được giải phóng và tái sử dụng nhiều lần để tổng hợp thêm một số lượng lớn acetyl-CoA từ pyruvic acid. Còn lại, toàn bộ phần acetyl được sử dụng để tạo nên phân tử citric acid. Trong các giai đoạn kế tiếp của chu trình citric acid, có rất nhiều phân tử nước tham gia vào quá trình, trong khi carbon dioxide và nguyên tử hydro được giải phóng ra ở các giai đoạn khác.
Kết quả cuối cùng của toàn bộ chu trình citric acid được giải thích, chứng minh rằng với mỗi phân tử glucose ban đầu sau khi được chuyển hóa ra được hai phân tử acetyl-CoA sẽ đi vào chu trình citric acid cùng với 6 phân tử nước. Những phân tử này sau đó sẽ thoái hóa thành 4 phân tử carbon dioxide, 16 nguyên tử hydro và 2 phân tử coenzyme A. Hai phân tử ATP được tạo thành theo cách thức mô tả dưới đây.
Sự hình thành ATP trong chu trình Citric Acid. Chu trình citric acid tự bản thân nó không giải phóng ra được nhiều năng lượng; chỉ một phân tử ATP được tạo thành trong duy nhất một phản ứng hóa học - trong quá trình từ α-ketoglutaric acid thành succinic acid. Do đó với mỗi phân tử glucose được chuyển hóa sẽ ra hai phân tử acetyl-CoA tham gia vào chu trình citric acid, mỗi phân tử chỉ tạo ra một phân tử ATP thành ra có tổng cộng là 2 phân tử ATP đã được hình thành.
Chức năng của enzym Dehydrogenases và coenzym Nicotinamide Adenine Dinucleotide trong việc giải phóng nguyên tử Hydro ở chu trình Citric Acid. Như đã nhấn mạnh tại một số điểm trong phần thảo luận này, nguyên tử hydro được giải phóng từ các phản ứng khác nhau trong chu trình citric acid - 4 nguyên tử hydro từ quá trình đường phân, 4 trong quá trình tổng hợp acetyl-CoA từ pyruvic acid, và 16 trong chu trình citric acid; do đó tổng cộng là 24 nguyên tử hydro được giải phóng từ phân tử glucose ban đầu. Mặc dù vậy, nguyên tử Hydro không tồn tại tự do trong dịch nội bào. Thay vào đó chúng được giải phóng ra từng đôi, và trong trường hợp này là nhờ xúc tác của một enzym đặc hiệu có tên là dehydrogenase. 20 trong tổng số 24 nguyên tử hydro ngay lập tức liên kết với nicotinamide adenine dinucleotide (NAD+), một dẫn xuất của vitamin B3 (niacin), theo phản ứng:
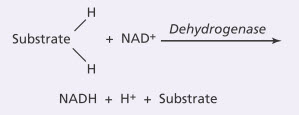
Phản ứng này sẽ không xảy ra nếu không có enzym dehydrogenase hoặc không có NAD+ làm nhiệm vụ như một chất mang hydro. Cả ion H+ tự do và hydro gắn với NAD+ sau đó đều tham gia vào các phản ứng oxy hóa để tổng hợp ra một số lượng lớn ATP, như đề cập sau đây.
4 nguyên tử hydro còn lại giải phóng từ quá trình thoái hóa của glucose - chính xác là trong chu trình citric acid ở giai đoạn chuyển từ succinic thành fumaric acid - liên kết với một enzym dehydrogenase đặc hiệu nhưng sau đó không được giải phóng vào NAD+. Thay vào đó chúng đi trực tiếp từ enzym dehydrogenase vào quá trình oxy hóa.
Chức năng của enzym Decarboxylases trong việc giải phóng carbon dioxide. Nhắc lại những phản ứng hóa học trong chu trình citric acid, cũng như quá trình hình thành acetyl-CoA từ pyruvic acid, chúng ta thấy rằng có 3 giai đoạn giải phóng ra carbon dioxide. Để giải phóng carbon dioxide, có những enzyme đặc hiệu gọi là decarboxylases, làm nhiệm vụ cắt carbon dioxide ra từ cơ chất. carbon dioxide sau đó được hoàn tan vào trong dịch của cơ thể và vận chuyển đến phổi, tại đó nó được đào thải ra khỏi cơ thể.
Bài viết cùng chuyên mục
Vai trò của O2 trong điều hòa hô hấp: điều hòa hô hấp bởi thụ thể ngoại vi
Oxygen không có ảnh hưởng trực tiếp tới trung tâm hô hấp của não trong việc điều hòa hô hấp. Thay vào đó, nó tác động gần như hoàn toàn lên các hóa thụ thể ở ngoại vi nằm trong động mạch cảnh và thân động mạch chủ.
Khuếch tán khí qua màng hô hấp: các yếu tố ảnh hưởng
Hệ số khuếch tán khí phụ thuộc vào khả năng hòa tan của khí trong màng tế bào, và hệ số này tỷ lệ nghịch với trọng lượng phân tử. Tỷ lệ khuếch tán khí trong màng hô hấp là gần như chính xác tương tự như trong nước.
Tăng vận chuyển ô xy đến mô: CO2 và H+ làm thay đổi phân ly oxy-hemoglobin (hiệu ứng bohr)
Khi máu đi qua các mô, CO2 khuếch tán từ tế bào ở mô vào máu, sự khuếch tán này làm tăng PCO2 máu, do đó làm tăng H2CO3 máu (axit cacbonic) và nồng độ ion H+. Hiệu ứng này sẽ làm chuyển dịch đồ thị phân ly oxy- hemoglobin sang bên phải và đi xuống.
Sự tiết Progesterone của nhau thai
Progesterone góp phần vào sự phát triển của thai, làm tăng bài tiết của ống dẫn trứng và tử cung người mẹ ddeer cung cấp chất dinh dưỡng thích hợp cho sự phát triển của phôi dâu và túi phôi.
Sự dẫn truyền cảm giác: đặc điểm trong con đường trước bên
Hệ trước bên là hệ thống dẫn truyền chưa phát triển bằng hệ thống cột tủy sau - dải cảm giác giữa. Thậm chí, các phương thức cảm giác nhất định chỉ được dẫn truyền trong hệ thống này.
Insulin kích hoạt receptor tế bào đích và những kết quả mang lại
Insulin liên kết với tiểu đơn vị của thụ thể của nó, gây ra quá trình tự phosphoryl hóa thụ thể - tiểu đơn vị, từ đó gây ra hoạt hóa tyrosine kinase.
Ngoại tâm thu nhĩ: rối loạn nhịp tim
Khi tim co sớm hơn bình thường, tâm thất chưa nhận đầy máu như bình thường và nhát bóp đó bơm ít máu hơn. Do đó sóng đập của nhát bóp đó lên thành mach sẽ yếu hơn thậm chí là yếu đến mức không thể bắt được gọi làm mạnh chìm.
Sóng vận mạch huyết áp: dao động của hệ thống điều chỉnh phản xạ huyết áp
Khoảng thời gian cho mỗi chu kỳ là 26 giây đối với chó đã gây mê, 7-10 giây ở người không gây mê. Sóng này được gọi là sóng vận mạch hay sóng Mayer.
Đám rối thần kinh cơ ruột và đám rối thần kinh dưới niêm mạc
Đám rối thần kinh cơ ruột không hoàn toàn có tác dụng kích thích vì một số neuron của nó có tác dụng ức chế; tận cùng của các sợi đó tiết ra một chất ức chế dẫn truyền, có thể là “polypeptide hoạt mạch ruột”.
Chức năng điều hoà nội môi sinh lý của thận
Áp suất thẩm thấu do các chất hòa tan trong huyết tương tạo nên, đặc biệt là Na+. Thông qua quá trình điều hòa thành phần các chất trong huyết tương, thận đã điều hòa áp suất thẩm thấu của dịch ngoại bào, bảo đảm áp suất này hằng định ở khoảng 300 mOsm/L.
Vòng phản xạ thần kinh: sự ổn định và mất ổn định
Hầu như tất cả các phần của não kết nối trực tiếp hoặc gián tiếp với tất cả các phần khác, nó tạo ra một thách thức nghiêm trọng. Nếu phần đầu tiên kích thích phần thứ hai, phần thứ hai kích thích phần thứ ba, phần thứ ba đến phần thứ tư và cứ như vậy.
Vòng phản xạ thần kinh: tín hiệu đầu ra liên tục
Các nơ-ron, giống các mô bị kích thích khác, phóng xung lặp đi lặp lại nếu mức điện thế màng kích thích tăng lên trên một ngưỡng nào đó. Điện thế màng của nhiều nơ-ron bình thường vẫn cao đủ để khiến chúng phóng xung liên tục.
Chức năng của Mineralocorticoids Aldosterone
Aldosterol chiếm phần lớn hoạt tính mineralocorticoid của hormon vỏ thượng thận, nhưng corticoid là glucocorticoid chính được tiết ở vỏ thượng thận, cũng tham giá đáng kể vào hoạt tính của mineralocorticoid.
Trạm thần kinh: sự chuyển tiếp các tín hiệu
Một số lượng lớn các tận cùng thần kinh từ mỗi sợi đến nằm trên các nơ-ron gần nhất trong “vùng” của nó, nhưng một số lượng nhỏ hơn các tận cùng thần kinh thì nằm trên các nơ-ron cách rất xa.
Vai trò của vùng dưới đồi điều hòa nhiệt độ cơ thể
Nhiệt độ của cơ thể được điều chỉnh hầu như hoàn toàn bởi cơ chế điều khiển thần kinh, và hầu hết mọi cơ chế này tác dụng thông qua trung tâm điều hòa nhiệt nằm ở vùng dưới đồi.
Bài tiết ion bicacbonat của tuyến tụy
Khi tuyến tụy bị kích thích để bài tiết ra một lượng dịch tụy dồi dào, nồng độ bicacbonat có thể lên cao tới khoảng 145 mEq/L, gấp khoảng 5 lần nồng độ ionbicacbonat trong huyết tương.
Các vùng các lớp và chức năng của vỏ não: cảm giác thân thể
Vùng cảm giác thân thể I nhạy cảm hơn và quan trọng hơn nhiều so với vùng cảm giác bản thể II đến mức trong thực tế, khái niệm “vỏ não cảm giác thân thể” hầu như luôn luôn có ý chỉ vùng I.
Sinh lý nơ ron thần kinh
Thân nơ ron có chức năng dinh dưỡng cho nơ ron. Ngoài ra, thân nơ ron có thể là nơi phát sinh xung động thần kinh và cũng có thể là nơi tiếp nhận xung động thần kinh từ nơi khác truyền đến nơ ron.
Giải phẫu và sinh lý của tụy
Tụy tiết nhiều hormon, như amylin, somatostatin, và pancreatic polypeptide, chức năng của chúng chưa được biết rõ. Mục đích chính là bàn về vai trò sinh lý của insulin và glucagon và sinh lý bệnh của các bệnh lý.
Thành phần của hệ renin angiotensin
Khi huyết áp động mạch giảm, phản ứng nội tại trong thận tạo ra nhiều phân tử prorenin trong các tế bào cận cầu thận để phân cắt và giải phóng renin.
Tác dụng của Insulin lên chuyển hóa chất béo
Insulin có nhiều tác dụng dẫn đến dự trữ chất béo tại mô mỡ. Đầu tiên, insulin tăng sử dụng glucose ở hầu hết các mô, điều này tự động làm giảm sử dụng chất béo, do đó, chức năng này như là dự trữ chất béo.
Sự phát triển của hệ cơ quan thai nhi
Sự phát triển các tế bào trên mỗi cơ quan thường chưa được hoàn thiện và cần 5 tháng mang thai còn lại để phát triển hoàn toàn. Ngay cả lúc sinh, những cấu trúc nhất định, đặc biệt là hệ thần kinh, thận và gan, thiếu sự phát triển hoàn toàn, như được mô tả sau.
Cơ chế tác động nội bào của testosterone
Testosteron kích thích tăng sản xuất protein ở hầu hết các mô trong cơ thể, đặc biệt là ở các mô cơ quan chịu trách nhiệm cho sự phát triển đặc tính sinh dục nam nguyên phát hoặc thứ phát.
Sự tiết estrogen của nhau thai
Estrogen chủ yếu gây ra một sự tăng sinh trên hầu hết các cơ quan sinh sản và liên quan đến người mẹ. Trong khi mang thai estrogen làm cho tử cung được mở rộng, phát triển vú và ống vú người mẹ, mở rộng cơ quan sinh dục ngoài.
Phosphate vô cơ ở dịch ngoại bào
Giá trị trung bình của tổng số lương phosphate đươc đưa ra khoảng 4 mg/dl, thay đổi giữa giá trị bình thường là 3 đến 4 mg/dl người lớn and 4 đến 5 mg/dl ở trẻ nhỏ.
