- Trang chủ
- Sách y học
- Sinh lý y học
- Cảm giác xúc giác: sự phát hiện và dẫn truyền
Cảm giác xúc giác: sự phát hiện và dẫn truyền
Mặc dù, cảm giác đụng chạm, áp lực và rung là phân loại thường gặp khi phân chia các cảm giác, nhưng chúng được nhận biết bởi các loại receptor giống nhau.
Biên tập viên: Trần Tiến Phong
Đánh giá: Trần Trà My, Trần Phương Phương
Mối liên hệ qua lại giữa các cảm giác xúc giác: đụng chạm, áp lực và rung
Mặc dù, cảm giác đụng chạm, áp lực và rung là phân loại thường gặp khi phân chia các cảm giác, nhưng chúng được nhận biết bởi các loại receptor giống nhau. Có 3 sự khác nhau cơ bản giữa chúng là: (1) cảm giác đụng chạm thường là kết quả của sự kích thích lên receptor xúc giác ở da hoặc trong mô ngay dưới da; (2) cảm giác áp lực thường có được từ sự biến dạng của các mô nằm sâu hơn; và (3) cảm giác rung là kết quả của những tín hiệu cảm giác lặp đi lặp lại nhanh chóng, nhưng một số loại receptor tương tự chúng cũng được dùng để nhận biết sự đụng chạm và áp lực.
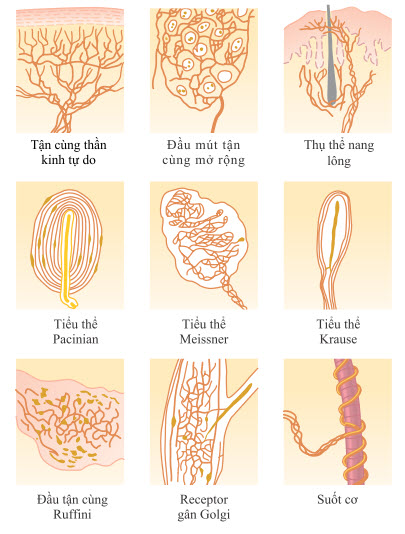
Hình. Một số loại kết thúc thần kinh cảm giác.
Các receptor xúc giác
Có ít nhất 6 loại receptor xúc giác khác nhau hoàn toàn, nhưng còn có nhiều loại khác tương tự chúng, các đặc điểm riêng biệt của chúng sẽ được nêu sau đây.
Đầu tiên, một số tận cùng thần kinh tự do, cái mà được tìm thấy ở mọi vùng trên da và nhiều mô khác, có thể phát hiện sự đụng chạm và áp lực. Ví dụ, mặc dù ánh sáng tiếp xúc với giác mạc mắt, nơi không chứa bất kỳ một loại tận cùng thần kinh nào khác ngoại trừ các tận cùng thần kinh tự do, nhưng nó có thể giúp nhận biết các cảm giác đụng chạm và áp lực.
Thứ hai, một receptor xúc giác rất nhạy cảm là tiểu thể Meissner một tận cùng thần kinh có vỏ bọc kéo dài (typ A beta) của sợi thần kinh cảm giác có myelin loại lớn. Bên trong vỏ bọc là rất nhiều sợi tơ thần kinh có đầu tận cùng phân nhánh. Các tiểu thể này có mặt ở những phần không có lông trên da và đặc biệt phong phú ở đầu ngón tay, môi và các vùng khác của da, những nơi có khả năng phân biệt vị trí không gian của cảm giác đụng chạm thì các tiểu thể này rất phát triển. Tiểu thể Meissner thích nghi trong chưa đến một giây sau khi chúng bị kích thích, nghĩa là chúng đặc biệt nhạy cảm với những chuyển động của vật thể qua bề mặt da, cũng như sự rung lắc có tần số thấp.
Thứ ba, các đầu ngón tay và các vùng chứa một số lượng lớn tiểu thể Meissner cũng chứa một lượng lớn các receptor xúc giác có đầu mút mở rộng, một dạng của đĩa Merkel. Các phần của lông trên da chứa một lượng trung bình các receptor có đầu mút mở rộng, mặc dù chúng không hoàn toàn là các tiểu thể Meissner. Các receptor này khác với tiểu thể Meissner ở chỗ ban đầu chúng dẫn một tín hiệu mạnh rồi đến tín hiệu thích nghi một phần, sau đó là tín hiệu tiếp tục yếu hơn tức là chúng thích nghi chậm. Do đó, chúng đảm nhiệm việc nhận biết các tín hiệu có trạng thái ổn định, cho phép con người xác định được những tiếp xúc liên tục của vật thể trên da.
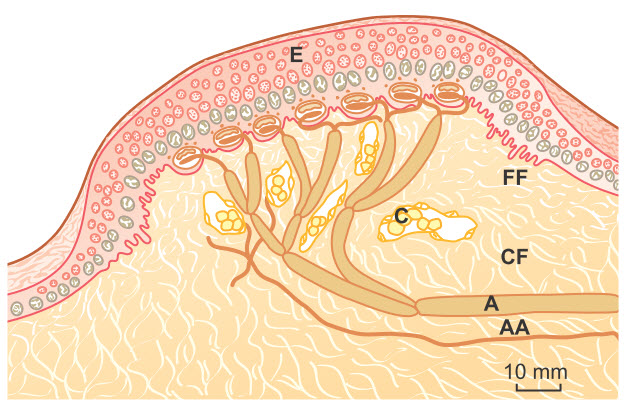
Hình. Một receptor hình vòm Iggo. Lưu ý số lượng nhiều đĩa Merkel kết nối với một sợi lớn có bao myelin (A) và tiếp giáp chặt chẽ với mặt dưới của biểu mô. AA, sợi trục không có myelin; C, ống mao dẫn; CF, các bó sợi collagen tự nhiên; E, lớp biểu bì dày lên của tiểu thể cảm ứng; FF, các bó sợi collagen nhỏ.
Các đĩa Merkel thường được nhóm thành một cơ quan nhận cảm gọi là receptor vòm Iggo, là phần lồi lên của lớp biểu bì da. Phần lồi lên này làm cho lớp biểu bì tại vị trí này nhô ra ngoài, từ đó tạo thành một hình vòm và chứa một số lượng lớn receptor nhạy cảm. Cũng cần chú ý rằng toàn bộ đĩa Merkel được phân bố bởi những sợi thần kinh đơn lớn có myelin (typ A beta). Các receptor này cùng với tiểu thể Meissner nêu trên đóng vai trò cực kỳ quan trọng trong việc khu trú cảm giác đụng chạm ở những vùng riêng biệt của cơ thể và trong việc xác định cấu tạo của thứ nó cảm nhận được.
Thứ tư, sự dịch chuyển nhẹ của vài sợi lông trên cơ thể kích thích một sợi thần kinh phân bố quanh chân lông. Như vậy, mỗi sợi lông và sợi thần kinh quanh chân lông được gọi là cơ quan chân lông, chúng cũng được gọi là các receptor đụng chạm. Một receptor thích nghi dễ dàng và, giống như tiểu thể Meissner, chủ yếu nhận biết (a) những chuyển động của vật thể trên bề mặt của cơ thể hoặc (b) mới tiếp xúc với cơ thể.
Thứ năm, nằm ở lớp sâu hơn của da và ở các mô nằm sâu bên trong cơ thể là nhiều tận cùng Ruffini, là loại receptor phân thành rất nhiều nhánh, có vỏ bọc. Những tận cùng này thích nghi rất chậm và do đó, nó quan trọng trong việc báo hiệu những trạng thái biến dạng liên tục của mô, như là sự đụng chạm mạnh kéo dài và tín hiệu áp lực. Chúng cũng được tìm thấy trong bao khớp và giúp báo hiệu mức độ xoay của khớp.
Thứ sáu, tiểu thể Pacinian nằm ngay ở dưới da và sâu trong cân mạc. Chúng chỉ bị kích thích bởi sự đè ép nhanh tại một vị trí trên mô vì chúng thích nghi chỉ trong vài phần trăm giây. Do đó, chúng đặc biệt quan trọng trong việc phát hiện sự rung lắc ở mô hoặc những thay đổi nhanh trong trạng thái cơ học của cơ thể.
Sự dẫn truyền các tín hiệu xúc giác ở sợi thần kinh ngoại vi
Hầu hết các receptor cảm giác chuyên biệt, như tiểu thể Meissner, receptor vòm Iggo, thụ thể nang lông, tiểu thể Pacinian và tận cùng Ruffini dẫn truyền các tín hiệu theo sợi thần kinh typ A beta, là sợi dẫn truyền với tốc độ từ 30-70 m/giây. Trái ngược lại, các receptor xúc giác là tận cùng thần kinh tự do thì dẫn truyền tín hiệu chủ yếu theo sợi có myelin typ A delta loại nhỏ với tốc độ chỉ 5-30 m/giây.
Một số tận cùng thần kinh tự do dẫn truyền qua sợi không có myelin typ C với tốc độ từ dưới 1m đến 2 m/ giây; những tận cùng thần kinh này dẫn truyền tín hiệu đến tủy sống và phần dưới thân não, có lẽ chủ yếu dẫn truyền cảm giác buồn.
Như vậy, nhiều loại tín hiệu cảm giác quan trọng - những cái giúp xác định vị trí chính xác trên da, ghi lại những mức cường độ hoặc những thay đổi nhanh chóng về cường độ tín hiệu cảm giác - tất cả được dẫn truyền theo nhiều loại sợi thần kinh cảm giác dẫn truyền nhanh. Trái ngược lại, những loại tín hiệu thô sơ như áp lực, cảm giác đụng chạm ít khu trú và đặc biệt cảm giác buồn được dẫn truyền bằng các sợi thần kinh rất nhỏ với tốc độ chậm hơn nhiều, chúng cần nhiều khoảng trống nhỏ trong các bó sợi thần kinh hơn các sợi nhanh.
Nhận biết sự rung
Tất cả các receptor xúc giác đều tham gia phát hiện cảm giác rung, mặc dù các receptor khác nhau phát hiện các tần số khác nhau của sự rung. Tiểu thể Pacinian có thể phát hiện tín hiệu rung trong khoảng từ 30-800 chu kỳ/giây do chúng đáp ứng cực nhanh với những biến dạng nhỏ và nhanh của mô. Chúng cũng dẫn truyền các tín hiệu qua sợi thần kinh typ A beta, là sợi có thể dẫn truyền nhiều đến 1000 xung mỗi giây. Trái ngược lại, sự rung với tần số thấp, từ 2-80 chu kỳ/giây, kích thích lên các receptor xúc giác khác, đặc biệt là tiểu thể Meissner, là loại thích nghi chậm hơn so với tiểu thể Pacinian.
Nhận biết cảm giác buồn và ngứa nhờ các tận cùng thần kinh cơ học
Các nghiên cứu về sinh lý thần kinh đã chứng minh sự tồn tại của các tận cùng thần kinh cơ học nhạy cảm và thích nghi nhanh, loại chỉ nhận cảm giác buồn và ngứa. Hơn nữa, các tận cùng thần kinh này hầu như chỉ phát hiện được ở lớp bề mặt của da, cũng là phần mô mà từ đó có thể gây ra cảm giác buồn và ngứa. Những cảm giác này được dẫn truyền bởi các sợi thần kinh rất nhỏ typ C, không có bao myelin, giống với những sợi dẫn truyền cảm giác đau, loại đau chậm.
Mục đích của cảm giác ngứa có lẽ là để thu hút sự chú ý với những kích thích bề mặt nhạy cảm như có một con bọ bò trên da hoặc một con ruồi sắp cắn và sau đó các tín hiệu gợi ý gây ra phản xạ gãi hoặc các hành động khác để loại bỏ vật chủ gây ra kích thích. Cảm giác ngứa có thể được giảm bớt bằng cách gãi nếu hành động này loại bỏ được tác nhân kích thích hoặc nếu gãi đủ mạnh để gây ra đau. Tín hiệu đau được tin rằng có thể chặn lại tín hiệu ngứa trong tủy bằng sự ức chế bên.
Bài viết cùng chuyên mục
Vai trò của các nhân não và tiền đình: nâng đỡ cơ thể chống lại trọng lực
Các nhân lưới được chia làm 2 nhóm chính: các nhân lưới ở cầu não, nằm ở phía sau bên của cầu não và kéo dài tới hành não, các nhân lưới ở hành não, kéo dài suốt toàn bộ hành não, nằm ở cạnh đường giữa.
Hoạt động chuyển hóa tế bào của Aldosterol
Aldosterol không có tác dụng vận chuyển natri ngay lập tức; đúng hơn tác dụng này phải chờ hình thành liên tục những chất đặc biệt cần thiết vận chuyển natri.
Biệt hóa tế bào cơ thể người
Trên thực tế, điện tử micrographs gợi ý rằng một số phân đoạn của vòng xoắn DNA được quấn xung quanh lõi histone trở nên rất đặc rằng họ không còn tháo dây đã cuốn để tạo thành các phân tử RNA.
Mức độ thiếu oxy mà một trẻ sơ sinh có thể chịu đựng được
Khi phế nang mở, hô hấp có thể bị ảnh hưởng thêm với vận động hô hấp tương đối yếu. May mắn thay, hít vào của trẻ bình thường rất giàu năng lượng; Có khả năng tạo ra áp lực âm trong khoang màng phổi lên đến 60mmHg.
Kiểm soát mạch máu bởi các ion và các yếu tố hóa học
Hầu hết các chất giãn mạch và co mạch đều có tác dụng nhỏ trên lưu lượng máu trừ khi chúng thay đổi tốc độ chuyển hóa của mô: trong hầu hết các trường hợp, lưu lượng máu tới mô và cung lượng tim không thay đổi.
Giải phóng hormon thyroxine và triiodothyronine từ tuyến giáp
Trong quá trình biến đổi phân tử thyroglobulin để giải phóng thyroxine và triiodothyronine, các tyrosine được iod hóa này cũng được giải thoát từ phân tử thyroglobulin, chúng không được bài tiết vào máu.
Xác định dung tích cặn chức năng, thể tích cặn, dung tích toàn phổi
Thiết bị đo dung tích với thể tích được làm đầy bởi không khí trộn với khí Heli. Trước khi thở từ thiết bị này, mỗi người được thở ra bình thường. Kết thúc thở ra, thể tích duy trì trong phổi bằng FRC.
Các giai đoạn bài tiết ở dạ dày
Bài tiết ở dạ dày ở giai đoạn kích thích tâm lý diễn ra thậm chí trước khi thức ăn đi vào dạ dày, đặc biệt khi đang được ăn. Sự bài tiết này là kết quả của quá trình trước khi thức ăn vào miệng như nhìn, ngửi, nghĩ hoặc nếm thức ăn.
Cung lượng tim: nghiên cứu định lượng
Tăng khả năng bơm máu cùng với tăng áp suất khoang màng phổi làm cung lượng tim đạt đỉnh vì tăng hoạt động tim nhưng đường cong lại dịch sang phải vì áp suất khoang màng phổi tăng.
Giảm chức năng thận: gây tăng huyết áp mãn tính
Mức độ tăng vừa phải của huyết áp cũng dẫn đến sự rút ngắn kỳ vọng sống. Tăng huyết áp nghiêm trọng nghĩa là giá trị huyết áp trung bình tăng 50% hoặc ở trên ngưỡng bình thường thì kỳ vọng sống là không lớn hơn một vài năm, trừ khi được điều trị thích hợp.
Sinh lý hệ thần kinh tự động
Receptor tiếp nhận norepinephrin của hệ giao cảm được gọi là noradrenergic receptor, bên cạnh norepinephrin, các receptor này cũng đáp ứng với epinephrin.
Ngừng tim trong rối loan nhịp tim
Giảm oxy máu khiến cho sợi cơ tim và sợi dẫn truyền mất điện thế màng bình thường và tính dễ kích thích này làm cho nhịp tự động biến mất.
Điện thế và thời gian chia chuẩn: điện tâm đồ bình thường
Những dòng dọc trên ECG là dòng thời gian chia chuẩn. Một ECG điển hình được chạy ở một tốc độ giấy 25mm/s, mặc dù tốc độ nhanh hơn đôi khi được sử dụng.
Phân ly ô xy hemoglobin thay đổi do BPG và lao động nặng
Trong khi lao động, một số yếu tố chuyển dịch đồ thị phân ly sang phải một cách đáng kể. Do đó cung cấp thêm O2 cho hoạt động, co cơ. Các cơ co sẽ giải phóng một lượng lớn khí CO2.
Trạm thần kinh: sự phân kỳ của các tín hiệu đi qua
Sự phân kỳ khuếch đại hiểu đơn giản là các tín hiệu đầu vào lan truyền đến một số lượng nơ-ron lớn hơn khi nó đi qua các cấp nơ-ron liên tiếp trong con đường của nó.
Cơ chế co cơ trơn
Một đặc tính quan trọng khác của cơ trơn, đặc biệt là loại cơ trơn đơn nhất nội tạng của nhiều cơ quan rỗng, là khả năng quay trở lại gần như lực co bóp ban đầu của nó vài giây hoặc vài phút sau khi nó bị kéo dài hoặc rút ngắn.
Các con đường gian nút và liên nhĩ: dẫn truyền xung động tim qua tâm nhĩ
Điện thế hoạt động bắt nguồn từ nút xoang đi ra ngoài vào trong các sợi cơ tâm nhĩ. Bằng cách này, điện thế hành động lan truyền qua toàn bộ khối cơ nhĩ và, cuối cùng, đến nút A-V.
Chức năng hành vi của vùng dưới đồi và cấu trúc liên kết với hệ limbic
Cùng với chức năng thực vật và nội tiết, sự kích thích hay thương tổn vùng dưới đồi cũng gây ảnh hưởng lớn đến hành vi cảm xúc của động vật và con người. Một số ảnh hưởng hành vi do sự kích thích vùng dưới đồi.
Phân loại cơ trơn
Cơ trơn ở mỗi cơ quan có các đặc điểm khác nhau: (1) kích thước (2) sự sắp xếp trong các bó (3) đáp ứng với các kích thích khác nhau (4) đặc điểm phân bố thần kinh (5) chức năng.
Đặc trưng của sự lan truyền tín hiệu trên thân dây thần kinh
Tốc độ lan truyền của điện thế hoạt động ở sợi thần kinh từ rất nhỏ 0,25 m / giây trong sợi không có myelin đến lớn như 100 m / giây (hơn chiều dài của một sân bóng đá trong 1 giây) trong sợi lớn có myelin.
Acid béo với alpha Glycerophosphate để tạo thành Triglycerides
Quá trình tổng hợp triglycerides, chỉ có khoảng 15% năng lượng ban đầu trong đường bị mất đi dưới dạng tạo nhiệt, còn lại 85% được chuyển sang dạng dự trữ triglycerides.
Phosphocreatine: kho lưu trữ năng lượng và như bộ đệm ATP
Phosphocreatine không thể hoạt động như một tác nhân trực tiếp vận chuyển năng lượng từ thức ăn đến các tế bào hoạt động chức năng, nhưng nó có thể vận chuyển năng lượng thông qua quá trình chuyển đổi với ATP.
Điều hòa bài tiết insulin
Kích thích tiết insulin bởi amino acid là quan trọng bởi vì insulin lần lượt tăng cường vận chuyển amino acid tới tế bào, cũng như sự hình thành protein trong tế bào..
Phức bộ QRS: hình dạng giãn rộng bất thường
Phức bộ QRS được xem là không bình thường khi kéo dài ít hơn 0,09s; khi nó giãn rộng trên 0,12s- tình trạng này chắc chắn gây ra bởi bệnh lý block ở 1 phần nào đó trong hệ thống dẫn truyền của tim.
Vận chuyển CO2 trong máu: các dạng vận chuyển và hiệu ứng Hanldane
Một lượng lớn CO2 đó có vai trò tạo nên sự cân bằng axit-bazơ của các chất dịch cơ thể. Dưới điều kiện bình thường khi nghỉ ngơi, trong mỗi 100 ml máu trung bình có 4ml CO2 được vận chuyển từ mô tới phổi.
